Содержание
Морские губки учат сопромату | Наука и жизнь
Некоторые морские организмы проявляют чудеса инженерной мысли, создавая удивительно прочные микроскопические структуры.
Инженеры подчас заимствуют идеи у самой природы. Существует даже такое направление в науке – бионика, которая изучает, как можно адаптировать природные технические решения для пользы человечеству. Например, берцовая кость человека, несмотря на свою изогнутую форму, может выдерживать вертикальную нагрузку больше тонны. Такую прочность ей обеспечивают особым образом ориентированные костные структуры. Похожий принцип был использован при проектировании Эйфелевой башни, в результате она и получилась такой легкой и прочной. Хотя наш инженер и архитектор Владимир Шухов использовал для своей знаменитой башни еще более легкую гиперболическую конструкцию, которую он сам и придумал. На единицу высоты Шуховской радиобашни было использовано в три раза меньше металла, чем у Эйфелевой башни в Париже.
Euplectella aspergillum – красивые морские губки, которые называют корзинками Венеры. Фото: NOAA Photo Library/Flickr.
Тонкими стеклянными нитями, спикулами, морские губки Euplectella aspergillum закрепляются на морском дне. От того, насколько крепко губка закрепится на поверхности, зависит без малого ее жизнь, поэтому естественный отбор наделил эти нити удивительной проч
Разрез спикулы морской губки (слева) и теоретическая модель материала с круговыми слоями(справа), которая обеспечивает максимальную прочность конструкции. Эволюция привела к возникновению биологической структуры, которая построена согласно правильным инже
‹
›
Открыть в полном размере
Но вернемся в мир природы, а именно к беспозвоночным. В отличие от тех, кто в ходе эволюции обзавёлся внутренним скелетам, вроде млекопитающих, птиц или рыб, беспозвоночным приходится, что называется, жить без «внутреннего стержня». Какие-то виды бесхребетное существование вполне устроило, другие же создали себе различные формы экзоскелета, чтобы лучше выживать во враждебном мире. Например, моллюски спрятались в прочные раковины, а крабы обзавелись не только крепким панцирем, но и грозными клешнями. Однако речь сейчас пойдёт не о них, а о другом интересном классе морских беспозвоночных – о стеклянных, или шестилучевых, губках. Самые известные, да и самые красивые представители их образуют семейство под названием корзинка Венеры.
Например, моллюски спрятались в прочные раковины, а крабы обзавелись не только крепким панцирем, но и грозными клешнями. Однако речь сейчас пойдёт не о них, а о другом интересном классе морских беспозвоночных – о стеклянных, или шестилучевых, губках. Самые известные, да и самые красивые представители их образуют семейство под названием корзинка Венеры.
Скелет этих губок представляет собой ажурный цилиндр, выполненный из переплетающихся волокон из диоксида кремния, или, грубо говоря, из стекла. Как оказалось, такие волокна по свойствам ничем не уступают современным оптоволоконным нитям. Вот только в промышленности они изготавливаются при температуре свыше 1000 градусов, в то время как губка, находясь на морском дне, спокойно делает продукт такого же качества при температуре всего лишь несколько градусов выше нуля. Действительно, производителям оптоволокна есть чему поучиться. Другая интересная особенность корзинки Венеры – то, как она крепится к морскому дну. Якорем ей служат длинные, толщиной с человеческий волос стеклянные нити, называемые базальными спикулами. На них находятся микроскопические крючки, благодаря которым губка прочно цепляется за поверхность. Казалось бы, использовать хрупкую стеклянную нить в качестве якорной цепи выглядит не самым продуманным шагом. Но это до тех пор, пока мы не посмотрим на структуру такой нити в разрезе.
На них находятся микроскопические крючки, благодаря которым губка прочно цепляется за поверхность. Казалось бы, использовать хрупкую стеклянную нить в качестве якорной цепи выглядит не самым продуманным шагом. Но это до тех пор, пока мы не посмотрим на структуру такой нити в разрезе.
Толщина «якорной» спикулы порядка 50 микрометров, что составляет одну двадцатую часть миллиметра. В центре нити располагается сердцевина, которую, в свою очередь, окружают несколько десятков концентрических слоев. Все слои и сердцевина сделаны из стекла, а ультратонкая прослойка между ними выполнена из органического материала. По виду такая картина очень напоминает срез ствола дерева. Фотография спикулы в разрезе так и оставалась бы иллюстрацией к какому-нибудь справочнику морфологии подводных обитателей, не попадись она на глаза профессору Ханешу Кесари с факультета инженерии Брауновского университета.И вот тут сработала так называемая профессиональная деформация взгляда на окружающий мир.
Если биологи видели всего лишь замысловатую концентрическую структуру, то инженер сразу же обратил внимание на толщину круговых слоев.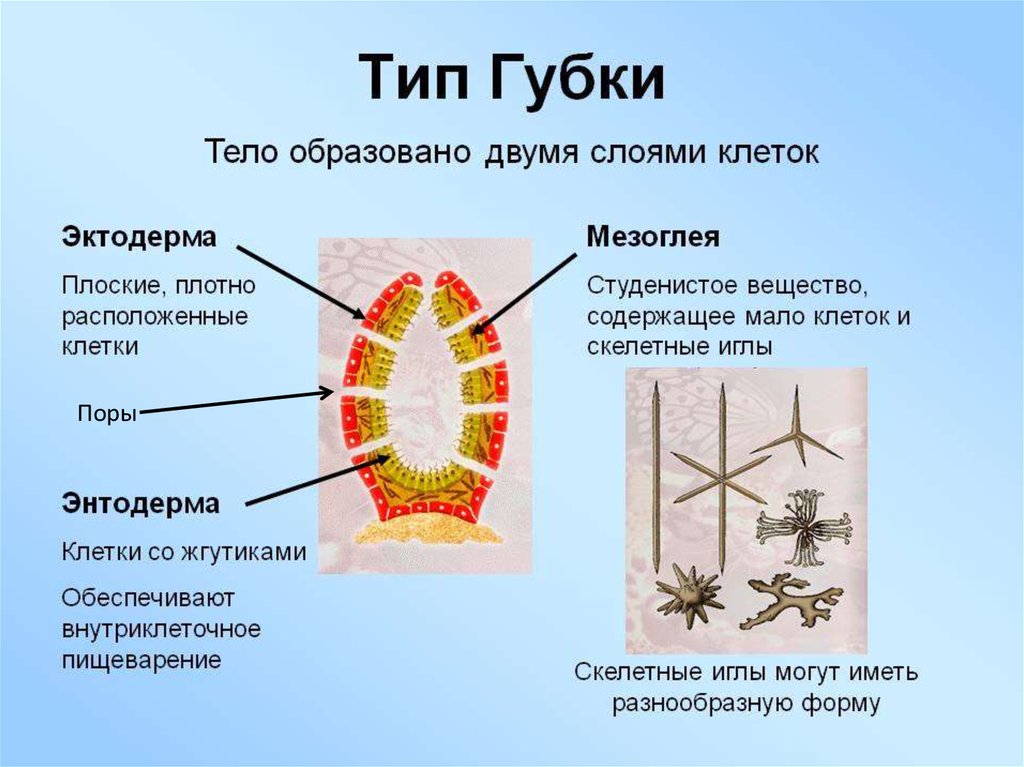 Она менялась от центра к наружному краю спикулы по вполне определенному закону: внутри были самые толстые слои, а по мере приближения к поверхности их толщина равномерно уменьшалась. И если морская губка в ходе эволюции использовала такое техническое решение для столь жизненно важной части тела, то, наверное, это было неспроста.
Она менялась от центра к наружному краю спикулы по вполне определенному закону: внутри были самые толстые слои, а по мере приближения к поверхности их толщина равномерно уменьшалась. И если морская губка в ходе эволюции использовала такое техническое решение для столь жизненно важной части тела, то, наверное, это было неспроста.
От того, насколько прочно губка закрепится на поверхности, зависит без малого ее жизнь. Поэтому прочность спикул, которыми она держится за дно, становится одним из важных факторов естественного отбора. Инженеры решили проверить, насколько строение стеклянных нитей губок соответствует наилучшей конструкции с точки зрения науки. Была создана модель, описывающая строение спикулы: сердцевина в виде сплошного стержня, окруженная концентрическими поверхностями переменной толщины. Задав параметры прочности материала, из которого губка изготавливает свои спикулы, исследователи рассчитали, какой должна быть оптимальная толщина слоев.
Удивительно, но результаты прочностных расчетов привели к тому же самому принципу, по которому губки конструируют свои «якори».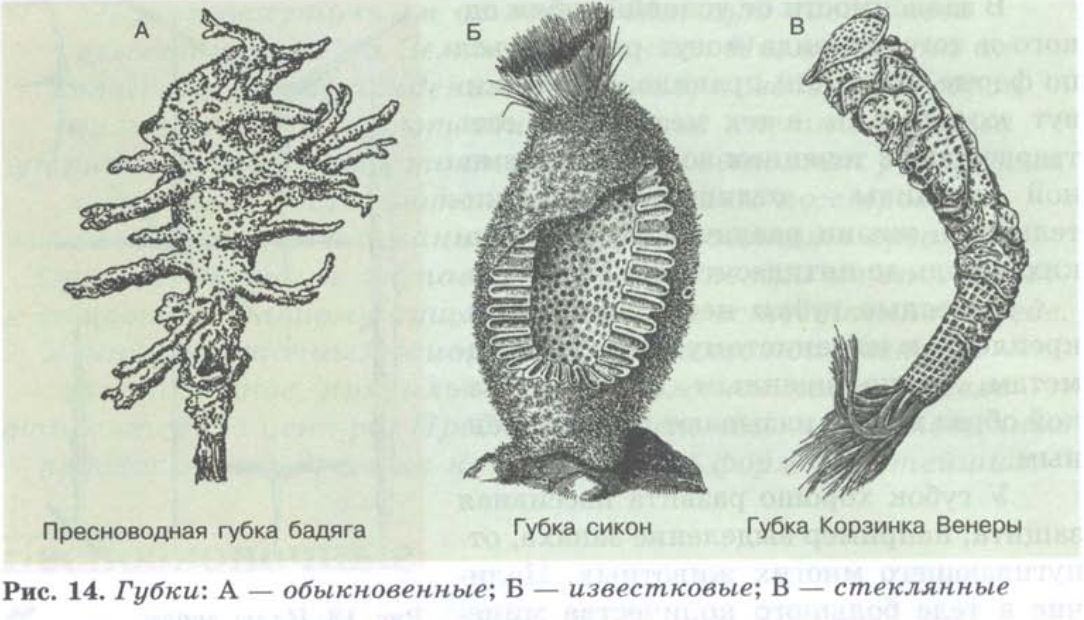 Профиль распределения толщины слоев теоретической модели получился в точности таким же, как в природе. Получается, что эволюция, не изучав сопромат в институте, пришла, тем не менее, к наиболее удачному инженерному решению.
Профиль распределения толщины слоев теоретической модели получился в точности таким же, как в природе. Получается, что эволюция, не изучав сопромат в институте, пришла, тем не менее, к наиболее удачному инженерному решению.
Фото: NOAA Photo Library/Flickr, Brown University
По материалам PNAS и Brown University
У губок не всё так просто, как кажется • Елена Наймарк • Новости науки на «Элементах» • Биология, Эволюция
Исследования последних лет показали, что самые простые многоклеточные — губки — представляют собой сложный симбиотический комплекс, включающий более сотни различных микроорганизмов. Многие из этих симбионтов встречаются только в губках. Более того, состав симбионтов у губок разных видов и в разных частях света оказался очень схожим. Это означает, что губки приобрели симбионтов в самом начале своей эволюции и по неизвестной причине сохранили их состав неизменным.
В последние 10–15 лет биологи начали пересматривать твердо установленные постулаты и перекраивать свою биологическую аксиоматику. Среди главных причин такого революционного движения — развитие новых методов микроскопирования и молекулярной биологии, в частности возможность читать генетические последовательности. Даже самый, казалось бы, простой организм — губка — в беспощадном свете новых методов видится уже довольно сложным.
Среди главных причин такого революционного движения — развитие новых методов микроскопирования и молекулярной биологии, в частности возможность читать генетические последовательности. Даже самый, казалось бы, простой организм — губка — в беспощадном свете новых методов видится уже довольно сложным.
Губку можно схематично представить в виде двух слоев клеток — эпителиальных и пищеварительных, а между слоями формируется твердый скелет из разного рода спикул (известковых или кремнезёмных игл). Хорошо известны старые опыты, в которых губку размалывали в кашу, а затем наблюдали, как клетки сами собой сортируются и снова складываются в полноценное животное. По сей день тянется дискуссия, следует ли относить губок к высокоорганизованным колониальным одноклеточным или к низкоорганизованным многоклеточным. Что может быть проще губок?
Но в последние годы выяснилось, что каждая губка — это конгломерат, включающий в себя более сотни (!) симбиотических организмов. Это стало очевидным после того, как исследователи из Алабамского университета в Бирмингеме (США) определили состав нуклеотидных последовательностей 16S-РНК у губок. Прежде симбионтов определяли с помощью лабораторного культивирования всей клеточной массы. Таким способом можно выявить только 5% микроорганизмов-симбионтов, потому что большинство симбионтов (да и свободноживущих микроорганизмов) в лабораторных пробирках не растут. Группу культивируемых в лаборатории микроорганизмов-симбионтов составили в основном фотосинтетики — цианобактерии и одноклеточные водоросли. Как и у кораллов, у губок эти микроорганизмы фиксируют углекислоту, снабжая гетеротрофную губку пищей и одновременно кислородом и энергией. Помимо них культивируются еще и актинобактерии.
Прежде симбионтов определяли с помощью лабораторного культивирования всей клеточной массы. Таким способом можно выявить только 5% микроорганизмов-симбионтов, потому что большинство симбионтов (да и свободноживущих микроорганизмов) в лабораторных пробирках не растут. Группу культивируемых в лаборатории микроорганизмов-симбионтов составили в основном фотосинтетики — цианобактерии и одноклеточные водоросли. Как и у кораллов, у губок эти микроорганизмы фиксируют углекислоту, снабжая гетеротрофную губку пищей и одновременно кислородом и энергией. Помимо них культивируются еще и актинобактерии.
Но кто еще, помимо фотосинтетиков, оказался сожителем губок? По 16S-РНК опознали архебактерий, альфапротеобактерий, актинобактерий, бактерий-нитрификаторов. В общей сложности микромир губок оказался весьма разнообразным: около 100 видов, принадлежащих к 14 типам бактерий, 2 типам архебактерий и нескольким видам эукариот. Многие из этих микроорганизмов определены пока только внутри губок, а в воде вокруг их нет. Вполне возможно, что своих симбионтов губка-родитель передает по наследству потомству: в сперматозоидах и в личинках губок обнаружили симбиотических бактерий. Замечательно, что набор симбионтов разных видов губок из разных частей света оказался очень сходным. Это означает, во-первых, что губки приобрели своих симбионтов в самом начале эволюционного пути. Во-вторых, что жизнеспособность и функционирование такого организма, как губка, требует совершенно определенного набора условий, который обеспечивается специфическим набором симбионтов.
Вполне возможно, что своих симбионтов губка-родитель передает по наследству потомству: в сперматозоидах и в личинках губок обнаружили симбиотических бактерий. Замечательно, что набор симбионтов разных видов губок из разных частей света оказался очень сходным. Это означает, во-первых, что губки приобрели своих симбионтов в самом начале эволюционного пути. Во-вторых, что жизнеспособность и функционирование такого организма, как губка, требует совершенно определенного набора условий, который обеспечивается специфическим набором симбионтов.
Получается, чтобы быть губкой, недостаточно иметь два слоя клеток и скелетные иглы, необходимо еще присутствие сотни действующих лиц. Какие из этих действующих лиц главные и обязательные, а какие второстепенные и необязательные — это ученым еще предстоит выяснить. Может ли вообще губка жить без своих симбионтов? Нужно понимать, что определить функции каждого из сожителей по отдельности практически невозможно — ведь их нельзя культивировать и, следовательно, выяснить, кто чем живет. Пока что удается лишь проследить с помощью радиоактивных меченых атомов путь отдельных веществ по цепочке микроорганизмов. Самих микроорганизмов «помечают» при этом разноцветными флуоресцентными белками. Так что можно видеть, что едят различные участники губкового консорциума.
Пока что удается лишь проследить с помощью радиоактивных меченых атомов путь отдельных веществ по цепочке микроорганизмов. Самих микроорганизмов «помечают» при этом разноцветными флуоресцентными белками. Так что можно видеть, что едят различные участники губкового консорциума.
Все эти исследования заставляют философски и методологически пересмотреть вопрос о сущности целостного организма. Даже такой просто устроенный организм, как губка, на самом деле оказался набором из множества различных организмов, каждый из которых предпочитает совместную жизнь единоличному обитанию. Другой вопрос: почему сами губки эволюционировали, а их сожители сохранили консервативный облик? Ведь у всех губок набор симбионтов более или менее единообразен. Всё это вопросы для нового поколения биологов.
Но исследователи убеждены, что их работа имеет не только академический интерес. Известно, что вытяжки губок обладают хорошим лечебным эффектом для многих заболеваний, в частности подавляют рост раковых клеток. Вполне возможно, что клиническое действие оказывают не сами губки, а их симбионты. Ведь очевидно, что одна из функций симбионтов — это защищать хозяина от нежелательных паразитов. В этом случае имеет смысл поискать среди бактерий тех, кто реально может справиться с паразитарными инфекциями. Так, вещество манзамин А (manzamine), выделенное из губок, гораздо действеннее в лечении малярии, чем все известные препараты. Ученым из Института биотехнологий Мэрилендского университета в Балтиморе (США) удалось показать, что это вещество синтезируется всеми видами губок, так что вероятность его бактериального происхождения чрезвычайно высока. Если ученые найдут эту гипотетическую бактерию, то наладить производство манзамина А при нынешних технологиях будет не очень сложно.
Вполне возможно, что клиническое действие оказывают не сами губки, а их симбионты. Ведь очевидно, что одна из функций симбионтов — это защищать хозяина от нежелательных паразитов. В этом случае имеет смысл поискать среди бактерий тех, кто реально может справиться с паразитарными инфекциями. Так, вещество манзамин А (manzamine), выделенное из губок, гораздо действеннее в лечении малярии, чем все известные препараты. Ученым из Института биотехнологий Мэрилендского университета в Балтиморе (США) удалось показать, что это вещество синтезируется всеми видами губок, так что вероятность его бактериального происхождения чрезвычайно высока. Если ученые найдут эту гипотетическую бактерию, то наладить производство манзамина А при нынешних технологиях будет не очень сложно.
Источник: Gretchen Vogel. The Inner Lives of Sponges // Science. 23 May 2008. V. 320. P. 1028–1030 (DOI: 10.1126/science.320.5879.1028).
Елена Наймарк
«Первичность губок» опережает по очкам «первичность гребневиков» • Сергей Ястребов • Новости науки на «Элементах» • Эволюция, Зоология
Продолжаются дискуссии вокруг вопроса о том, какая ветвь многоклеточных животных является самой древней: губки или гребневики? На данный момент похоже, что губки «побеждают». Но это, в свою очередь, порождает новые не решенные пока вопросы, важнейший из которых касается происхождения нервной системы.
Но это, в свою очередь, порождает новые не решенные пока вопросы, важнейший из которых касается происхождения нервной системы.
В поисках предка
Человек вышел из животного царства. В ходе своей истории он постоянно так или иначе имел дело с другими животными: собаки, лошади, киты, акулы, жирафы, страусы, жуки, дождевые черви, скорпионы, морские звезды — все они были частью мира, знакомого людям с глубокой древности. Животными при этом считали всех, кто не растения. Правда, натуралистов издавна озадачивали живые существа, сочетавшие (на первый взгляд) признаки животных и растений и относившиеся невесть куда — например, актинии или губки. Но таких существ было относительно немного — во всяком случае, заметных невооруженным глазом. На протяжении тысячелетий, от Аристотеля до Декарта, общая картина виделась более-менее стабильной и ясной.
Настоящий переворот в представлениях о структуре живой природы произошел после великой научной революции XVII века. В 1677 году в лондонском журнале Philosophical Transactions of the Royal Society (который продолжает выходить по сей день) была опубликована знаменитая работа Антони ван Левенгука «О мельчайших животных».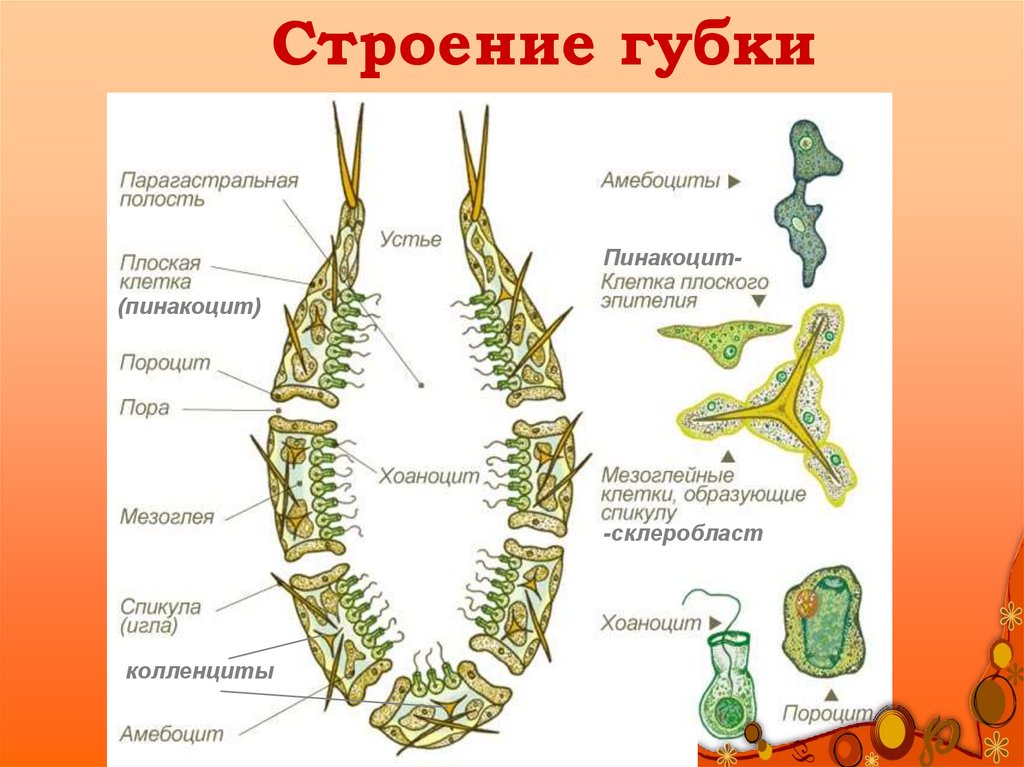 Перед людьми начал приоткрываться огромный, разнообразный, ни на что не похожий мир микроскопических организмов. Сначала этот мир казался исследователям совершенно беспорядочным. Кое-какую ясность внесла провозглашенная в 1839 году Теодором Шванном клеточная теория, согласно которой и животные, и растения состоят из того или иного числа клеток, в целом подобных одна другой. Отсюда логически следовало, что некоторые организмы могут состоять и из одной клетки. И действительно, вышедшее в 1848 году «Руководство по сравнительной анатомии» Карла фон Зибольда и Германа Станниуса открывается утверждением, что все животные делятся на одноклеточных и многоклеточных (автором этого важнейшего обобщения был Зибольд, который написал том, посвященный беспозвоночным). Теперь стало понятно, что микроскопические живые существа — во всяком случае, подавляющее большинство их — это просто-напросто одноклеточные.
Перед людьми начал приоткрываться огромный, разнообразный, ни на что не похожий мир микроскопических организмов. Сначала этот мир казался исследователям совершенно беспорядочным. Кое-какую ясность внесла провозглашенная в 1839 году Теодором Шванном клеточная теория, согласно которой и животные, и растения состоят из того или иного числа клеток, в целом подобных одна другой. Отсюда логически следовало, что некоторые организмы могут состоять и из одной клетки. И действительно, вышедшее в 1848 году «Руководство по сравнительной анатомии» Карла фон Зибольда и Германа Станниуса открывается утверждением, что все животные делятся на одноклеточных и многоклеточных (автором этого важнейшего обобщения был Зибольд, который написал том, посвященный беспозвоночным). Теперь стало понятно, что микроскопические живые существа — во всяком случае, подавляющее большинство их — это просто-напросто одноклеточные.
Только после работ Левенгука, Шванна и Зибольда стала возможной (а потом и необходимой) постановка проблемы происхождения многоклеточных животных. Надо заметить, что в современной биологии «животными» вообще принято называть только многоклеточных существ (Metazoa), никакие одноклеточные в эту группу не входят, так что словосочетание «многоклеточные животные» — теперь лишь дань традиции. Но в любом случае ясно, что у многоклеточных животных должны были быть одноклеточные предки. Что это за предки? Как произошел переход между уровнями организации — с одноклеточного на многоклеточный? Как выглядели переходные формы? Эти вопросы, впервые осознанные в середине XIX века, занимают биологов до сих пор.
Надо заметить, что в современной биологии «животными» вообще принято называть только многоклеточных существ (Metazoa), никакие одноклеточные в эту группу не входят, так что словосочетание «многоклеточные животные» — теперь лишь дань традиции. Но в любом случае ясно, что у многоклеточных животных должны были быть одноклеточные предки. Что это за предки? Как произошел переход между уровнями организации — с одноклеточного на многоклеточный? Как выглядели переходные формы? Эти вопросы, впервые осознанные в середине XIX века, занимают биологов до сих пор.
Проще всего ответить на первый вопрос — о том, кем были предки. Ближайшие родственники животных известны: это воротничковые жгутиконосцы (рис. 2). В терминах современной эволюционной биологии многоклеточные животные и воротничковые жгутиконосцы — сестринские группы. Близкое родство воротничковых жгутиконосцев с животными предположили еще в XIX веке, сразу после открытия этих микроскопических организмов, а сейчас оно надежно подтверждается молекулярной генетикой (см. Расшифрован геном хоанофлагеллят — ближайших одноклеточных родичей всех многоклеточных животных, «Элементы», 18.02.2008). Эти группы имеют огромное количество общих генов, включая такие, которые раньше считались уникальным для многоклеточных животных «ноу-хау» (например, у воротничковых жгутиконосцев нашлись гены, кодирующие кадгерины и интегрины — белки, обычно участвующие во взаимодействии животных клеток между собой). Насчет их родства сомнений нет.
Расшифрован геном хоанофлагеллят — ближайших одноклеточных родичей всех многоклеточных животных, «Элементы», 18.02.2008). Эти группы имеют огромное количество общих генов, включая такие, которые раньше считались уникальным для многоклеточных животных «ноу-хау» (например, у воротничковых жгутиконосцев нашлись гены, кодирующие кадгерины и интегрины — белки, обычно участвующие во взаимодействии животных клеток между собой). Насчет их родства сомнений нет.
Многоклеточные животные, воротничковые жгутиконосцы и еще несколько групп одноклеточных или колониальных организмов образуют большую группу, которая называется Holozoa. В свою очередь, группа Holozoa входит в супергруппу Opisthokonta (кроме голозой, то есть животных и их родственников, к опистоконтам относятся грибы и их родственники). Опистоконты, или заднежгутиковые, — это одна из примерно десятка крупнейших эволюционных ветвей эукариот, организмов со сложной ядросодержащей клеткой (см. Случайно открытый жгутиконосец обновляет систему эукариот, «Элементы», 06. 02.2019). Таково место животных на общем эволюционном древе.
02.2019). Таково место животных на общем эволюционном древе.
Итак, воротничковые жгутиконосцы — ближайшие современные родственники многоклеточных животных. Значит ли это, что они и есть их предки? Нет, не обязательно. Во-первых, надо помнить, что сколь угодно близкое родство между двумя живыми существами само по себе еще не означает, что одно из них произошло от другого (например, человек не является ни предком, ни потомком своих ближайших современных родственников — шимпанзе и бонобо). Во-вторых, у современных воротничковых жгутиконосцев есть признаки, противоречащие гипотезе, что многоклеточные животные произошли непосредственно от них. Например, сравнительная генетика совершенно однозначно показывает, что у общего предка всей супергруппы Opisthokonta был ген, который называется Brachyury (см. У одноклеточных организмов есть ген, способный управлять развитием хорды, «Элементы», 25.10.2013). Для многоклеточных животных этот ген очень важен: он регулирует ранние стадии их эмбрионального развития и, в частности, закладку хорды у позвоночных. Есть он и у многих неживотных опистоконт, причем не только среди голозой, но и в другой эволюционной ветви, ведущей к грибам. Более того, ген Brachyury широко распространен не только у колониальных, но и у одноклеточных опистоконт (вероятно, его продукт как-то регулирует взаимное расположение клеток при делении). Но вот у воротничковых жгутиконосцев его нет. Они его утратили. Между тем многоклеточные животные, конечно, могли произойти только от таких предков, у которых ген Brachyury еще был — иначе как бы они его унаследовали? Значит, если и они и произошли от воротничковых, то не от современных.
Есть он и у многих неживотных опистоконт, причем не только среди голозой, но и в другой эволюционной ветви, ведущей к грибам. Более того, ген Brachyury широко распространен не только у колониальных, но и у одноклеточных опистоконт (вероятно, его продукт как-то регулирует взаимное расположение клеток при делении). Но вот у воротничковых жгутиконосцев его нет. Они его утратили. Между тем многоклеточные животные, конечно, могли произойти только от таких предков, у которых ген Brachyury еще был — иначе как бы они его унаследовали? Значит, если и они и произошли от воротничковых, то не от современных.
Еще один повод для размышления дает физиологически обусловленная синхронность делений клеток (в этом случае общее число клеточных ядер в организме какое-то время остается степенью двойки: 2, 4, 8, 16…). Такая синхронность наблюдается при дроблении оплодотворенного яйца большинства многоклеточных животных и — с другой стороны — при образовании колонии у некоторых голозой, не относящихся к воротничковым жгутиконосцам (см.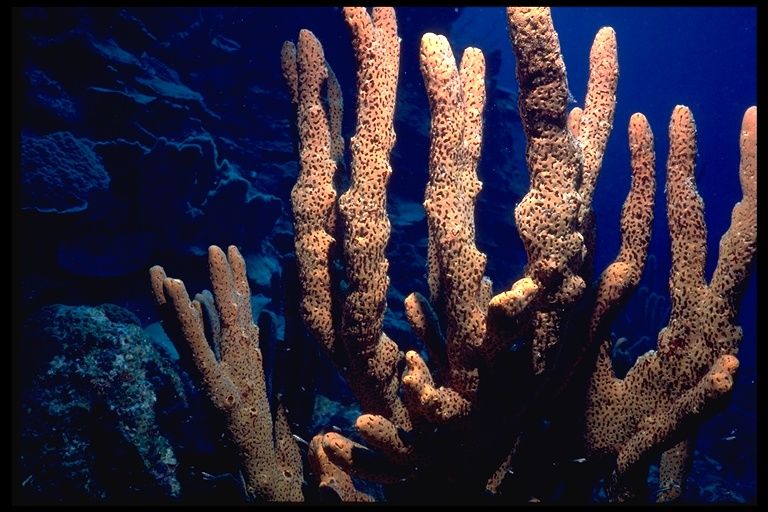 Ядра мезомицетозоев делятся синхронно, как у зародышей животных, «Элементы», 05.06.2013). У самих воротничковых ее нет. Все это, вероятно, означает, что жизненный цикл общего предка воротничковых жгутиконосцев и многоклеточных животных был гораздо более сложным, чем жизненный цикл современных воротничковых (который вторично упрощен). Непосредственный предок многоклеточных животных до нашего времени не дожил. Что и неудивительно: ведь с тех пор прошло, вероятно, около 600 миллионов лет.
Ядра мезомицетозоев делятся синхронно, как у зародышей животных, «Элементы», 05.06.2013). У самих воротничковых ее нет. Все это, вероятно, означает, что жизненный цикл общего предка воротничковых жгутиконосцев и многоклеточных животных был гораздо более сложным, чем жизненный цикл современных воротничковых (который вторично упрощен). Непосредственный предок многоклеточных животных до нашего времени не дожил. Что и неудивительно: ведь с тех пор прошло, вероятно, около 600 миллионов лет.
Гастрея
vs фагоцителла
Проблема происхождения многоклеточных животных — одна из самых старых проблем эволюционной биологии. За полтора века на эту тему были высказаны десятки гипотез, в том числе детально разработанных, которые принято повышать в статусе и называть теориями (см. А. В. Иванов, 1968. Происхождение многоклеточных животных). Многие из них сейчас устарели настолько, что представляют интерес только с точки зрения истории науки. Другие, может быть, частично и верны, но по разным причинам непопулярны. Главными соперниками с 1880-х годов и до сих пор выступают две теории (именно их в первую очередь вспомнит любой зоолог). Это теория гастреи Эрнста Геккеля и теория фагоцителлы Ильи Ильича Мечникова.
Главными соперниками с 1880-х годов и до сих пор выступают две теории (именно их в первую очередь вспомнит любой зоолог). Это теория гастреи Эрнста Геккеля и теория фагоцителлы Ильи Ильича Мечникова.
Теория Геккеля основана на том, что многоклеточным животным свойственны эмбриональные стадии бластулы и гаструлы. В варианте, который принято считать наиболее типичным, бластула представляет собой однослойный шар из жгутиковых клеток. Потом одно полушарие втягивается внутрь, и однослойный шар трансформируется в двуслойный мешок — гаструлу. Возникающая при этом внутренняя полость гаструлы (архентерон) становится кишечной полостью взрослого животного. Геккель предположил, что эти эмбриональные стадии повторяют (рекапитулируют) ход эволюционных событий. Вначале была шарообразная колония жгутиконосцев, похожая на бластулу, но представляющая собой взрослый организм — Геккель назвал ее бластеей. Переднее полушарие бластеи (преимущественно двигательное) так и осталось снаружи, а заднее (преимущественно пищеварительное) втянулось внутрь, образовав камеру со входом. Эта камера и стала первичной кишкой, или архентероном. Получившийся двуслойный организм Геккель назвал гастреей. Предполагается, что гастрея была подобна гаструле, но являлась не промежуточной стадией развития, а конечной (взрослой).
Эта камера и стала первичной кишкой, или архентероном. Получившийся двуслойный организм Геккель назвал гастреей. Предполагается, что гастрея была подобна гаструле, но являлась не промежуточной стадией развития, а конечной (взрослой).
Геккелевская гаструла, устроенная поразительно единообразно, образует своего рода «узел сходства» в индивидуальном развитии великого множества многоклеточных животных, от коралловых полипов до ланцетника включительно. Неудивительно, что Геккель посчитал широкую распространенность этой эмбриональной стадии следом общего для всех животных эволюционного прошлого. Образование гаструлы путем упорядоченного впячивания целого клеточного слоя называется инвагинацией. Типичную инвагинационную гаструлу можно увидеть у представителей самых разных типов животных, как бы сильно ни отличались между собой их взрослые формы (рис. 3).
Для Геккеля было важно, что инвагинационная гаструла преобладает у таких примитивных животных, как коралловые полипы (к которым относятся актинии). С другой стороны, этот способ развития не является всеобщим. Например, у гидроидных полипов (к которым относится пресноводная гидра) гаструляция чаще всего идет не путем инвагинации, а путем иммиграции: клетки будущей кишки по одной уходят в глубину зародыша. Нет инвагинационной гаструлы и у бескишечных ресничных червей, очень простых и примитивных представителей гигантской ветви двусторонне-симметричных животных (билатерий). Эти факты и привели Мечникова к мысли, что у потомков бластеи клетки первым делом начали уползать внутрь поодиночке, чтобы заниматься внутриклеточным пищеварением. В результате возникла фагоцителла — организм, состоящий из слоя движущих жгутиковых клеток снаружи и рыхлой массы переваривающих клеток (фагоцитов) внутри. Инвагинационная гаструла, по Мечникову, появилась гораздо позже — после того, как масса фагоцитов превратилась в кишечник, имеющий собственную полость (рис. 4).
С другой стороны, этот способ развития не является всеобщим. Например, у гидроидных полипов (к которым относится пресноводная гидра) гаструляция чаще всего идет не путем инвагинации, а путем иммиграции: клетки будущей кишки по одной уходят в глубину зародыша. Нет инвагинационной гаструлы и у бескишечных ресничных червей, очень простых и примитивных представителей гигантской ветви двусторонне-симметричных животных (билатерий). Эти факты и привели Мечникова к мысли, что у потомков бластеи клетки первым делом начали уползать внутрь поодиночке, чтобы заниматься внутриклеточным пищеварением. В результате возникла фагоцителла — организм, состоящий из слоя движущих жгутиковых клеток снаружи и рыхлой массы переваривающих клеток (фагоцитов) внутри. Инвагинационная гаструла, по Мечникову, появилась гораздо позже — после того, как масса фагоцитов превратилась в кишечник, имеющий собственную полость (рис. 4).
Веским доводом за теорию фагоцителлы и против теории гастреи было убеждение многих зоологов, что гидроидные полипы древнее и примитивнее коралловых. Однако это убеждение никогда не было всеобщим, а появившаяся в 1990-х годах молекулярная систематика животных сразу показала, что оно неверно (D. Bridge et al., 1992. Class-level relationships in the phylum Cnidaria: evidence from mitochondrial genome structure). Именно кораллы — самые древние и примитивные из современных стрекающих, а гидроиды — группа относительно молодая (см. Двусторонняя симметрия кораллов древнее радиальной, «Элементы», 26.10.2016). В этом отношении Геккель был прав.
Однако это убеждение никогда не было всеобщим, а появившаяся в 1990-х годах молекулярная систематика животных сразу показала, что оно неверно (D. Bridge et al., 1992. Class-level relationships in the phylum Cnidaria: evidence from mitochondrial genome structure). Именно кораллы — самые древние и примитивные из современных стрекающих, а гидроиды — группа относительно молодая (см. Двусторонняя симметрия кораллов древнее радиальной, «Элементы», 26.10.2016). В этом отношении Геккель был прав.
Сколько бы ни спорили между собой сторонники теорий гастреи и фагоцителлы, общего между этими теориями в любом случае гораздо больше, чем различий. Обе теории (по крайней мере в их классических версиях) предполагают, что исходной формой многоклеточных животных была шарообразная жгутиковая бластея, весь жизненный цикл которой проходил в толще морской воды. Такой жизненный цикл называется голопелагическим.
Плавающие шарообразные колонии, похожие на геккелевскую бластею, действительно существуют. Они независимо возникли у далеких друг от друга эукариот, относящихся к разным супергруппам: у золотистых водорослей (см. Synura), у зеленых водорослей (см. Volvox) и у воротничковых жгутиконосцев (см. Sphaeroeca). Последние, как мы знаем, и есть ближайшие родственники животных. Причем сфероэка, бластееподобный представитель воротничковых, была открыта немецким биологом Робертом Лаутерборном больше чем через двадцать лет после того, как Геккель описал гипотетическую бластею. Казалось бы, вот пример удачного предсказания! Но тут есть оговорки. Дело в том, что подавляющее большинство воротничковых жгутиконосцев питается не в плавающем, а в сидячем состоянии: к этому способу жизни они приспособлены гораздо лучше. Для них наиболее типичны прикрепленные колонии, имеющие форму лент, пластинок, звездочек или ветвящихся деревьев. Активно плавающая колония у них редкое исключение.
Они независимо возникли у далеких друг от друга эукариот, относящихся к разным супергруппам: у золотистых водорослей (см. Synura), у зеленых водорослей (см. Volvox) и у воротничковых жгутиконосцев (см. Sphaeroeca). Последние, как мы знаем, и есть ближайшие родственники животных. Причем сфероэка, бластееподобный представитель воротничковых, была открыта немецким биологом Робертом Лаутерборном больше чем через двадцать лет после того, как Геккель описал гипотетическую бластею. Казалось бы, вот пример удачного предсказания! Но тут есть оговорки. Дело в том, что подавляющее большинство воротничковых жгутиконосцев питается не в плавающем, а в сидячем состоянии: к этому способу жизни они приспособлены гораздо лучше. Для них наиболее типичны прикрепленные колонии, имеющие форму лент, пластинок, звездочек или ветвящихся деревьев. Активно плавающая колония у них редкое исключение.
Кроме того, с теорией гастреи Геккеля плохо согласуются губки — прикрепленные существа, которых многие исследователи считали (и считают до сих пор) самыми примитивными из всех многоклеточных животных. Геккель несколько преувеличил сходство ранних стадий развития губок, с одной стороны, и остальных многоклеточных животных, с другой (всех животных, кроме губок, иногда называют настоящими многоклеточными животными, Eumetazoa). У губок безусловно есть истинная бластула, причем у многих видов свободно плавающая — иными словами, бластула у них может быть личинкой. Но вот наличие у них истинной гаструлы спорно. Встречается обоснованное мнение, что ее просто нет (см., например: L. Salvini-Plawen, 1998. Morphologie: Haeckels gastraea-theorie und ihre folgen). А отсюда следует, что губки, вероятнее всего, отделились от других животных на эволюционной стадии бластеи (L. Salvini-Plawen, 1978. On the origin and evolution of the lower Metazoa). Стадию гастреи они не проходили.
Геккель несколько преувеличил сходство ранних стадий развития губок, с одной стороны, и остальных многоклеточных животных, с другой (всех животных, кроме губок, иногда называют настоящими многоклеточными животными, Eumetazoa). У губок безусловно есть истинная бластула, причем у многих видов свободно плавающая — иными словами, бластула у них может быть личинкой. Но вот наличие у них истинной гаструлы спорно. Встречается обоснованное мнение, что ее просто нет (см., например: L. Salvini-Plawen, 1998. Morphologie: Haeckels gastraea-theorie und ihre folgen). А отсюда следует, что губки, вероятнее всего, отделились от других животных на эволюционной стадии бластеи (L. Salvini-Plawen, 1978. On the origin and evolution of the lower Metazoa). Стадию гастреи они не проходили.
Могли ли настоящие многоклеточные животные, снабженные мускулатурой и нервной системой, произойти от неподвижных губок? И если могли, то как?
Вершки и корешки
Мы знаем, что многоклеточные животные и воротничковые жгутиконосцы относятся к эволюционной ветви Holozoa, входящей, в свою очередь, в состав супергруппы Opisthokonta. Название Opisthokonta — заднежгутиковые — связано с уникальным признаком этой группы: жгутиковые клетки опистоконт, в отличие от всех остальных эукариот, всегда движутся жгутиком назад. Откуда этот признак взялся? Скорее всего, дело в том, что одноклеточные предки заднежгутиковых питались в сидячем состоянии, и жгутик у них бился таким образом, чтобы создавать ток воды, поднимающий пищевые частицы с поверхности дна (В. В. Малахов, 2011. Современные представления о происхождении многоклеточных животных). При переходе к плаванию такой способ работы жгутика как раз и означает, что клетка будет двигаться жгутиком назад. Именно так плавают сперматозоиды животных и жгутиковые споры хитридиевых грибов (напомним, что грибы тоже опистоконты, они гораздо ближе к животным, чем, скажем, растения или инфузории). Древние опистоконты легко прикреплялись к грунту, потому что были в основном обитателями твердых поверхностей — дна или почвы, а не открытого водного пространства, как многие эукариоты из других супергрупп (например, одноклеточные зеленые водоросли, см.
Название Opisthokonta — заднежгутиковые — связано с уникальным признаком этой группы: жгутиковые клетки опистоконт, в отличие от всех остальных эукариот, всегда движутся жгутиком назад. Откуда этот признак взялся? Скорее всего, дело в том, что одноклеточные предки заднежгутиковых питались в сидячем состоянии, и жгутик у них бился таким образом, чтобы создавать ток воды, поднимающий пищевые частицы с поверхности дна (В. В. Малахов, 2011. Современные представления о происхождении многоклеточных животных). При переходе к плаванию такой способ работы жгутика как раз и означает, что клетка будет двигаться жгутиком назад. Именно так плавают сперматозоиды животных и жгутиковые споры хитридиевых грибов (напомним, что грибы тоже опистоконты, они гораздо ближе к животным, чем, скажем, растения или инфузории). Древние опистоконты легко прикреплялись к грунту, потому что были в основном обитателями твердых поверхностей — дна или почвы, а не открытого водного пространства, как многие эукариоты из других супергрупп (например, одноклеточные зеленые водоросли, см. Хламидомонада).
Хламидомонада).
Со жгутиком опистоконт связана еще одна проблема. Дело в том, что любой эукариотный жгутик закреплен в теле клетки с помощью так называемого корешкового аппарата, который устроен у разных эукариот совершенно по-разному (см. С. А. Карпов, 2001. Строение клетки протистов). Есть два главных типа корешковых аппаратов: ленты микротрубочек и исчерченные корешки, состоящие из микрофиламентов (микрофиламенты тоньше микротрубочек, но зато их пучки, в отличие от пучков микротрубочек, могут сокращаться). Электронно-микроскопические исследования, в ходе которых были открыты эти структуры, сразу показали, что тип корешкового аппарата часто служит важным систематическим признаком (см., например: С. А. Карпов, 1990. Система протистов).
Проблема же заключается вот в чем. У воротничковых жгутиконосцев корешковый аппарат состоит из лент микротрубочек, радиально расходящихся от основания единственного жгутика (поскольку клетка у воротничковых радиально-симметричная). У жгутиковых клеток губок — хоаноцитов, которые во всех отношениях очень похожи на воротничковых жгутиконосцев, корешковый аппарат тоже состоит из лент микротрубочек. А вот у всех остальных многоклеточных животных, от кораллов и плоских червей до человека, корешковые аппараты жгутиков и ресничек — это исчерченные корешки.
А вот у всех остальных многоклеточных животных, от кораллов и плоских червей до человека, корешковые аппараты жгутиков и ресничек — это исчерченные корешки.
Кроме того, и у воротничковых жгутиконосцев, и у хоаноцитов губок жгутик несет многочисленные тонкие придатки — мастигонемы (особенность, для нефотосинтезирующих эукариот очень редкая). У настоящих многоклеточных животных мастигонем не бывает никогда.
Тонкие особенности устройства живых клеток, для изучения которых нужен электронный микроскоп, в биологии принято называть ультраструктурой. Итак, между воротничковыми жгутиконосцами и хоаноцитами губок есть ультраструктурное сходство, которое свидетельствует о родстве воротничковых жгутиконосцев и губок друг с другом, но отличает и тех и других от настоящих многоклеточных животных. В 1980-х годах это послужило основанием для гипотезы, что от воротничковых жгутиконосцев произошли только губки, в то время как происхождение всех остальных многоклеточных животных — совершенно иное (Л. Н. Серавин, 1986. Природа и происхождение губок). Губок с воротничковыми выделяли в особое царство Parazoa (см. О. Г. Кусакин, А. Л. Дроздов, 1994. Филема органического мира. Ч. 1). От кого в таком случае произошли настоящие многоклеточные животные, оставалось загадкой. На роль их вероятных предков предлагались то хитридиевые грибы, то жгутиковые зеленые водоросли. На самом деле, как мы сейчас понимаем, хитридиевые грибы относятся к другой ветви опистоконт, а зеленые водоросли — вообще к другой супергруппе эукариот.
Н. Серавин, 1986. Природа и происхождение губок). Губок с воротничковыми выделяли в особое царство Parazoa (см. О. Г. Кусакин, А. Л. Дроздов, 1994. Филема органического мира. Ч. 1). От кого в таком случае произошли настоящие многоклеточные животные, оставалось загадкой. На роль их вероятных предков предлагались то хитридиевые грибы, то жгутиковые зеленые водоросли. На самом деле, как мы сейчас понимаем, хитридиевые грибы относятся к другой ветви опистоконт, а зеленые водоросли — вообще к другой супергруппе эукариот.
У настоящих многоклеточных животных тоже встречаются воротничковые клетки, напоминающие о воротничковых жгутиконосцах. Воротничок, которому воротничковые жгутиконосцы обязаны своим названием — это прозрачная воронка, окружающая жгутик и состоящая из множества отдельных микроворсинок, длинных тонких выростов с микрофиламентами внутри (см. рис. 1, А). У хоаноцитов губок он устроен очень похоже. Но эта структура не уникальна. Жгутиковые клетки с настоящим воротничком из микроворсинок описаны, например, у личинок некоторых полипов, причем как коралловых, так и гидроидных (K. M. Lyons, 1973. Evolutionary implications of collar cell ectoderm in a coral planula). Специальные исследования показали, что воротничковые клетки широко распространены у иглокожих (не только в покровах, но и в выстилке воднососудистой системы; см. рис. 5 и статью: A. Norrevang, K. G. Wingstrand, 1970. On the occurrence and structure of choanocyte-like cells in some echinoderms). Типичные воротничковые клетки найдены и в покровах полухордовых (A. Norrevang, 1964. Choanocytes in the skin of Harrimania kupfferi (Enteropneusta)), и в предротовой воронке ланцетника (T. Stach, 1996. On the preoral pit of the larval amphioxus (Branchiostoma lanceolatum)).
M. Lyons, 1973. Evolutionary implications of collar cell ectoderm in a coral planula). Специальные исследования показали, что воротничковые клетки широко распространены у иглокожих (не только в покровах, но и в выстилке воднососудистой системы; см. рис. 5 и статью: A. Norrevang, K. G. Wingstrand, 1970. On the occurrence and structure of choanocyte-like cells in some echinoderms). Типичные воротничковые клетки найдены и в покровах полухордовых (A. Norrevang, 1964. Choanocytes in the skin of Harrimania kupfferi (Enteropneusta)), и в предротовой воронке ланцетника (T. Stach, 1996. On the preoral pit of the larval amphioxus (Branchiostoma lanceolatum)).
Более того, преемственность воротничковых клеток дотянулась и до позвоночных. Именно от воротничковых клеток, судя по всему, произошли сенсорные клетки внутреннего уха, каждая из которых несет жгутик и группу микроворсинок (правда, жгутик там расположен не посреди микроворсинок, а эксцентрично). Недавно было показано, что существует большая группа общих генов, которые активны в сенсорных клетках внутреннего уха позвоночных и в хоаноцитах губок (J. F. Pena et al., 2016. Conserved expression of vertebrate microvillar gene homologs in choanocytes of freshwater sponges). Функции белков, кодируемых этими генами, связаны или со сборкой микрофиламентов, или с контактами микроворсинок между собой. Пример такого белка — виллин, который назван в честь микроворсинок (по-латыни microvilli) и регулирует сборку пучков микрофиламентов, дающих микроворсинкам опору. Виллин есть у настоящих многоклеточных животных, у губок, у воротничковых жгутиконосцев и у амебоидных Holozoa (см. Capsaspora), но не найден ни у каких других эукариот.
F. Pena et al., 2016. Conserved expression of vertebrate microvillar gene homologs in choanocytes of freshwater sponges). Функции белков, кодируемых этими генами, связаны или со сборкой микрофиламентов, или с контактами микроворсинок между собой. Пример такого белка — виллин, который назван в честь микроворсинок (по-латыни microvilli) и регулирует сборку пучков микрофиламентов, дающих микроворсинкам опору. Виллин есть у настоящих многоклеточных животных, у губок, у воротничковых жгутиконосцев и у амебоидных Holozoa (см. Capsaspora), но не найден ни у каких других эукариот.
Однако во всех жгутиковых клетках настоящих многоклеточных животных корешковый аппарат жгутика представлен исчерченным корешком, а не лентами микротрубочек. В этом отношении жгутиковые клетки настоящих многоклеточных животных (неважно, воротничковые или нет) резко отличаются и от воротничковых жгутиконосцев, и от хоаноцитов губок. Не разобравшись в причинах этих отличий, делать выводы о происхождении всех многоклеточных животных было нельзя.
Неотенические личинки губок?
К началу XXI века тщательные электронно-микроскопические исследования прояснили ситуацию. Оказалось, что у бластулообразных личинок губок жгутиковые клетки как раз могут обладать типичными исчерченными корешками, которые исчезают и сменяются лентами микротрубочек только при метаморфозе (см. M. Maldonado, 2004. Choanoflagellates, choanocytes, and animal multicellularity). Это установлено как минимум для нескольких видов губок, относящихся к разным классам. У жгутиков личинок губок есть и другие черты сходства со жгутиками настоящих многоклеточных животных: кое-какие детали расположения микротрубочек внутри жгутика, отсутствие воротничка и мастигонем (рис. 6).
Кроме того, жгутиковые клетки личинок губок нередко бывают подостланы базальной мембраной, которая характерна для эпителиев настоящих многоклеточных животных, а среди взрослых губок встречается только у представителей небольшого класса гомосклерид. У личинок тех же гомосклеридных губок описаны и десмосомы — межклеточные контакты весьма продвинутого типа, которые обычно обеспечивают в эпителиях настоящих многоклеточных животных механическое сцепление клеток друг с другом.
Строго говоря, эти факты — не такая уж и новость. Часть их была известна уже в шестидесятых-семидесятых годах XX века. Австрийский зоолог Луитфрид Сальвини-Плавен считал, что смена корешкового аппарата у губок связана с характером нагрузки: жгутикам плавающей личинки, в отличие от жгутиков неподвижной взрослой губки, надо обеспечивать поступательное движение животного в толще воды, потому они и снабжены корешками из микрофиламентов, способными активно сокращаться (см. L. Salvini-Plawen, 1978. On the origin and evolution of the lower Metazoa).
Получается, что никакой пропасти между губками и настоящими многоклеточными животными на самом деле нет.
Теперь подведем некоторый итог. У жгутиковых клеток настоящих многоклеточных животных для «заякоривания» жгутика в теле клетки служит исчерченный корешок, воротничка из микроворсинок вокруг жгутика чаще все же нет (несмотря на вышеперечисленные примеры, обязательным приложением к жгутику он тут не является), мастигонем не бывает, а поверхностный слой клеток почти всегда представляет собой полноценный эпителий с десмосомами и базальной мембраной. И мы видим, что по сумме структурных признаков настоящие многоклеточные животные гораздо более похожи на личинок губок, чем на взрослых губок.
И мы видим, что по сумме структурных признаков настоящие многоклеточные животные гораздо более похожи на личинок губок, чем на взрослых губок.
Вряд ли это случайность. В роли предка настоящих многоклеточных животных трудно представить взрослую губку, но достаточно легко — бластулообразную личинку губки. Напрашивается идея, что именно личинка губки и была этим предком. Конечно, речь идет о личинке, отказавшейся от метаморфоза и научившейся самостоятельно размножаться — явление, хорошо знакомое зоологам еще с XIX века и называемое неотенией (см., например: B. A. Pierce, H. M. Smith, 1979. Neoteny or paedogenesis?). Датский зоолог Клаус Нильсен так и назвал одну свою обзорную статью: «Шесть главных шагов в эволюции животных: являемся ли мы производными личинок губок?» (C. Nielsen, 2008. Six major steps in animal evolution: are we derived sponge larvae?) — и ответ он дает положительный: да, являемся.
Неотеническая личинка губки и есть геккелевская бластея. Во всяком случае, это возможно. Что касается дальнейшей эволюции Eumetazoa, то она вполне могла идти примерно так, как описывает геккелевская теория гастреи — но только с дополнением, касающимся происхождения исходной формы, бластеи. Если эта гипотеза верна, значит, мы все — прямые потомки неотенических личинок губок (рис. 7).
Что касается дальнейшей эволюции Eumetazoa, то она вполне могла идти примерно так, как описывает геккелевская теория гастреи — но только с дополнением, касающимся происхождения исходной формы, бластеи. Если эта гипотеза верна, значит, мы все — прямые потомки неотенических личинок губок (рис. 7).
К сожалению, надежно определимые элементы минерального скелета губок (они называются спикулами) появляются в палеонтологической летописи только в кембрийском периоде, то есть примерно тогда же, когда и все основные группы настоящих многоклеточных животных (см. А. Ю. Журавлев, 2014. Ранняя эволюция Metazoa — взгляд палеонтолога). Поэтому происхождение Eumetazoa от кого-то из современных классов губок маловероятно. Доводы в пользу того, что губки появились намного раньше, остаются большей частью косвенными (например, химические биомаркеры, которые, по некоторым предположениям, могут принадлежать и водорослям). Правда, надо учитывать, что у некоторых губок твердого скелета вообще нет (см. Oscarellidae).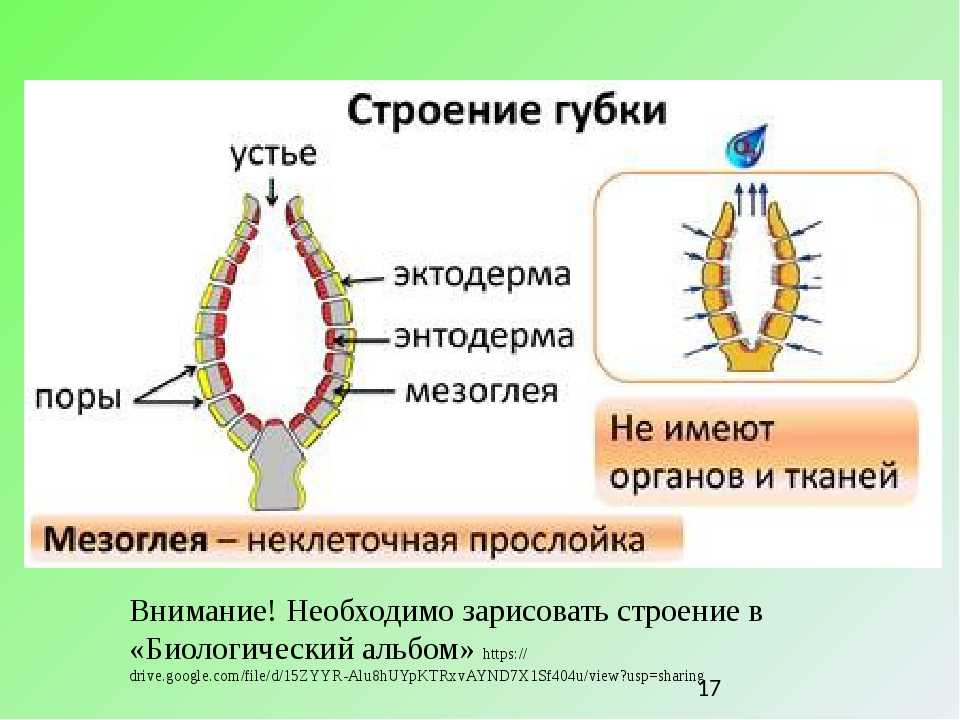 А если древнейшие губки были бесскелетными, то шансов сохраниться они имели немного.
А если древнейшие губки были бесскелетными, то шансов сохраниться они имели немного.
В 2015 году международная группа палеонтологов описала открытые в известном китайском местонахождения Доушаньтуо остатки древнего организма, который получил название Eocyathispongia qiania (Z. Yin et al., 2015. Sponge grade body fossil with cellular resolution dating 60 Myr before the Cambrian). Это было сидячее существо очень небольшого размера — около 2 миллиметров, и оно сохранилось достаточно хорошо, чтобы под электронным микроскопом можно было рассмотреть минерализованные остатки отдельных клеток. Общая форма тела, множество дырочек (видимо, служивших для входа воды), внутренние камеры, трубчатое устье, характерные очертания покровных клеток-пинакоцитов — все эти детали позволили палеонтологам предположить, что эоциатиспонгия была самой настоящей губкой. Правда, маленькой и бесскелетной. А жила она 600 миллионов лет назад, почти за 60 миллионов лет до начала кембрийского периода и до знаменитого кембрийского взрыва (см. С. А. Ястребов, 2016. Кембрийский взрыв). Первые бесспорные остатки настоящих многоклеточных животных появляются в летописи не более чем за 13–15 миллионов лет до начала кембрия (см. Кимберелла). Так что если эоциатиспонгия интерпретирована верно — значит, губки все-таки опередили настоящих многоклеточных животных примерно на 45 миллионов лет.
С. А. Ястребов, 2016. Кембрийский взрыв). Первые бесспорные остатки настоящих многоклеточных животных появляются в летописи не более чем за 13–15 миллионов лет до начала кембрия (см. Кимберелла). Так что если эоциатиспонгия интерпретирована верно — значит, губки все-таки опередили настоящих многоклеточных животных примерно на 45 миллионов лет.
Проблема гребневиков
Ситуация едва успела проясниться, как появилась новая гипотеза, основательно спутавшая карты классическим зоологам: самая древняя эволюционная ветвь многоклеточных животных — не губки, а гребневики. Эта проблема обсуждалась на страницах «Элементов» уже несколько раз, поэтому сейчас будет достаточно напомнить основные моменты. Гребневики — это полупрозрачные морские хищники, очень отдаленно похожие на медуз; подавляющее большинство их проводит всю свою жизнь, активно плавая в толще воды, но несколько представителей перешли к ползанию по дну (скорее всего, эволюционно недавно). Гипотеза «первичности гребневиков» впервые была высказана молекулярными биологами в 2013 году (см. Геном гребневиков говорит в пользу двукратного возникновения нервной системы у животных, «Элементы», 19.12.2013). Отсюда сразу же вытекала серьезная проблема, связанная с происхождением нервной системы. Дело в том, что у губок нервной системы нет и, судя по всему, никогда не было. Она есть, с одной стороны, у гребневиков, а с другой — у стрекающих и у двусторонне-симметричных, или билатерий (эти две группы образуют единую ветвь). Если верна гипотеза «первичности гребневиков», значит, ветви гребневиков и стрекающих с билатериями находятся на эволюционном древе слишком далеко друг от друга, чтобы их общий предок мог иметь нервную систему. Она должна была появиться независимо.
Геном гребневиков говорит в пользу двукратного возникновения нервной системы у животных, «Элементы», 19.12.2013). Отсюда сразу же вытекала серьезная проблема, связанная с происхождением нервной системы. Дело в том, что у губок нервной системы нет и, судя по всему, никогда не было. Она есть, с одной стороны, у гребневиков, а с другой — у стрекающих и у двусторонне-симметричных, или билатерий (эти две группы образуют единую ветвь). Если верна гипотеза «первичности гребневиков», значит, ветви гребневиков и стрекающих с билатериями находятся на эволюционном древе слишком далеко друг от друга, чтобы их общий предок мог иметь нервную систему. Она должна была появиться независимо.
Вскоре гипотеза «первичности гребневиков» получила новые подтверждения, причем как со стороны исследования родственных связей, так и со стороны генов, функции которых связаны с работой нервной системы и мышц (см. Гипотеза о двукратном появлении нервной системы получила новые подтверждения, «Элементы», 26.05.2014). По сравнению со стрекающими и билатериями гребневики имеют совершенно иной набор нейромедиаторов, рецепторов, синаптических белков и некоторых других молекул, обладающих сигнальными функциями. Гены семейства Hox, играющие огромную роль в дифференцировке отделов тела (включая нервную систему) и у билатерий, и у стрекающих, у гребневиков отсутствуют вообще. Архитектура нервной системы — причем на всех возможных уровнях — у них альтернативная. В этом отношении гребневики еще больше похожи на инопланетян, чем иглокожие, облик которых дал инопланетным разумным существам Питер Уоттс в знаменитом романе «Ложная слепота».
По сравнению со стрекающими и билатериями гребневики имеют совершенно иной набор нейромедиаторов, рецепторов, синаптических белков и некоторых других молекул, обладающих сигнальными функциями. Гены семейства Hox, играющие огромную роль в дифференцировке отделов тела (включая нервную систему) и у билатерий, и у стрекающих, у гребневиков отсутствуют вообще. Архитектура нервной системы — причем на всех возможных уровнях — у них альтернативная. В этом отношении гребневики еще больше похожи на инопланетян, чем иглокожие, облик которых дал инопланетным разумным существам Питер Уоттс в знаменитом романе «Ложная слепота».
Все это выглядит достаточно убедительно. Однако еще одна серьезная проблема заключается вот в чем: после принятия гипотезы «первичности гребневиков» становится очень трудно представить, как мог бы выглядеть общий предок всех животных (если, конечно, не считать его одноклеточным). Стройный сценарий происхождения всех настоящих многоклеточных животных от губок разрушается — ведь губки теперь оказываются более молодой группой, чем подвижные, хищные, имеющие нервную систему гребневики, — а взамен ничего толком не предлагается.
В 2015 году группа известных зоологов, в которую входил тот же Клаус Нильсен, опубликовала большую статью с анализом этой проблемы (см. Дискуссия о роли гребневиков в эволюции продолжается, «Элементы», 18.09.2015). Эти авторы подчеркнули, что вопрос о положении гребневиков на эволюционном древе и вопрос о происхождении нервной системы суть в общем случае разные вопросы, которые лучше бы разделять, чтобы не получилась путаница. Что касается положения гребневиков на древе, то тут есть многочисленные техническое сложности, связанные с анализом генных последовательностей этой группы животных — а она в любом случае очень древняя, и в ее истории были бурные эпизоды вроде почти полного вымирания в момент мел-палеогенового кризиса, после которого, однако, разнообразие гребневиков быстро восстановилось; неудивительно, что запутаннные генетические следы таких эволюционных событий сбивают биоинформатиков с толку. Самостоятельно решать вопрос о родственных связях гребневиков Нильсен и его коллеги не пытаются. Происхождение нервной системы — другое дело. Нейробиологические данные (сами по себе не связанные с темой родства) убедительно показывают, что нервная система гребневиков устроена крайне своеобразно, если сопоставлять ее с нервной системой стрекающих и билатерий. Вопрос в том, с какого уровня они все стартовали.
Происхождение нервной системы — другое дело. Нейробиологические данные (сами по себе не связанные с темой родства) убедительно показывают, что нервная система гребневиков устроена крайне своеобразно, если сопоставлять ее с нервной системой стрекающих и билатерий. Вопрос в том, с какого уровня они все стартовали.
Между тем молекулярные филогенетики, занимающиеся именно проблемой родственных связей между разными группами животных, продолжали свои исследования. В том же 2015 году появилась скрупулезно выполненная работа, в которой сравнивалась надежность гипотез «первичности гребневиков» и «первичности губок» с точки зрения топологии эволюционного древа (Сравнительная геномика вынуждает пересмотреть место гребневиков на эволюционном древе животных, «Элементы», 18.12.2015). Было показано, что решение этой задачи неустойчиво: оно зависит от выбранной вычислительной методики и от того, какие группы организмов, помимо животных, включены в анализ (например, в одном случае для сравнения используются данные по грибам, в другом по воротничковым жгутиконосцам — и выводы получаются разными). В общем, результат здесь в любом случае на грани достоверности. Если же аккуратно сопоставить все возможные методики и сочетания выбранных групп, то чаша весов, похоже, склоняется всё-таки в сторону «первичности губок».
В общем, результат здесь в любом случае на грани достоверности. Если же аккуратно сопоставить все возможные методики и сочетания выбранных групп, то чаша весов, похоже, склоняется всё-таки в сторону «первичности губок».
Мнение Нильсена
Несколько недель назад вышел очередной обзор всего этого комплекса проблем, на сей раз подписанный Клаусом Нильсеном как единственным автором. Знаменитый датский зоолог суммирует накопившиеся факты и предлагает эволюционный сценарий, который кажется ему самым вероятным.
Прежде всего, за последние годы вышло еще несколько молекулярно-филогенетических работ, где на основе постоянно растущих наборов данных вновь и вновь отстаивается гипотеза «первичности губок» (см., например, рис. 8 и статью: P. Simion et al., 2017. A large and consistent phylogenomic dataset supports sponges as the sister group to all other animals). Гипотеза «первичности гребневиков» там объявляется вычислительным артефактом, научной ошибкой, и делается вывод, что гребневики — только вторая по порядку отхождения ветвь эволюционного древа животных (после губок).
Конечно, это не означает немедленного прекращения дискуссий по данному вопросу. Не исключено, что окончательное «укоренение» ветвей губок и гребневиков вообще находится за пределом разрешающей способности молекулярной филогенетики как метода (W. Pett et al., 2019. The role of homology and orthology in the phylogenomic analysis of metazoan gene content). Такое мнение тоже надо принять во внимание. Похожую проблему можно наблюдать, например, в лингвистике (см. Лингвистическая эволюция сходна с биологической, «Элементы», 18.10.2007). Языковеды прекрасно справляются с установлением родственных связей языков внутри семей (скажем, индоевропейских языков друг с другом), а вот поиск родственных связей между разными языковыми семьями — так называемая макрокомпаративистика — остается хотя и интересной, но крайне дискуссионной темой (см. Г. С. Старостин, 2016. К истокам языкового разнообразия). Наряду с энтузиастами макрокомпаративистики есть исследователи, считающие, что сколько-нибудь надежные результаты в этой области невозможны принципиально, потому что для языков, разошедшихся более 8–10 тысяч лет назад, следы родства становятся неотличимы от случайных совпадений. Анализ генетических текстов, конечно, позволяет проникать на несравненно большую глубину (вплоть до миллиардов лет), но его разрешающая способность тоже не бесконечна: рано или поздно следы родства забиваются «шумом», и с этим ничего сделать уже нельзя.
Анализ генетических текстов, конечно, позволяет проникать на несравненно большую глубину (вплоть до миллиардов лет), но его разрешающая способность тоже не бесконечна: рано или поздно следы родства забиваются «шумом», и с этим ничего сделать уже нельзя.
В любом случае гипотезу «первичности гребневиков» сейчас никак нельзя признать победившей, и к гипотезе «первичности губок» приходится относиться серьезно. Как рабочая версия она до сих пор пригодна.
Что же теперь можно сказать о происхождении нервной системы? Характерно, что авторы вышеупомянутых статей о «первичности губок» — молекулярные филогенетики, занимающиеся проблемами родства — решать этот вопрос не берутся (во всяком случае, пока), а оставляют открытыми несколько возможностей.
Всего есть пять эволюционных ветвей многоклеточных животных: губки, гребневики, пластинчатые, стрекающие и билатерии. Самая маленькая ветвь — пластинчатые: всего два известных представителя, трихоплакс и описанная в 2018 году хойлунгия, отличающаяся от трихоплакса исключительно генетически. Самая огромная ветвь — билатерии: около полутора миллионов описанных видов (две трети из которых — насекомые). У гребневиков, стрекающих и билатерий нервная система есть, у губок и у пластинчатых ее нет. Это сухие факты, а вот дальше начинаются интерпретации.
Самая огромная ветвь — билатерии: около полутора миллионов описанных видов (две трети из которых — насекомые). У гребневиков, стрекающих и билатерий нервная система есть, у губок и у пластинчатых ее нет. Это сухие факты, а вот дальше начинаются интерпретации.
Существует гипотеза, что нервная система когда-то была у всех животных, но губки и пластинчатые ее по каким-то причинам утратили (см., например: J. F. Ryan, M. Chiodin, 2015. Where is my mind? How sponges and placozoans may have lost neural cell types). Нильсен прямо называет эту гипотезу плодом воображения. Нервная система — штука весьма универсальная («адаптация общего значения» в терминах Ивана Ивановича Шмальгаузена), она может пригодиться почти что при любом образе жизни; несколько случаев ее потери у животных действительно известно, но все они связаны с переходом к паразитизму, а ни для губок, ни для пластинчатых нет оснований такое предполагать. Сильным аргументом в пользу большой древности нервной системы часто считают наличие у губок и у пластинчатых ряда генов, которые ассоциированы с нервной системой у ее обладателей (продукты этих генов обеспечивают срабатывание ионных каналов, выброс из клетки нейромедиатора и тому подобное). Однако сами по себе эти генетические факты мало что доказывают. Ген всего лишь определяет структуру белка, а физиологические эффекты этого белка в ходе эволюции могут меняться, и иногда очень сильно.
Однако сами по себе эти генетические факты мало что доказывают. Ген всего лишь определяет структуру белка, а физиологические эффекты этого белка в ходе эволюции могут меняться, и иногда очень сильно.
Например, есть такое понятие, как протосинаптические белки (P. Burkhardt, S. G. Sprecher, 2017. Evolutionary origin of synapses and neurons — bridging the gap). Синапс — это специфический контакт между нервными клетками, через который передается сигнал (химический или электрический). Клетки, образующие синапс, оснащены в зоне контакта сложными молекулярными конструкциями, в создании которых участвуют десятки белков, кодируемых соответствующими генами. Многие из этих синаптических белков есть не только у животных с нервной системой, но и у губок. На первый взгляд это хорошее основание для гипотезы, что у губок раньше были нервные клетки. Но значительная доля синаптических белков обнаружена (благодаря исследованиям геномов и транскриптомов) не только у губок, но и у воротничковых жгутиконосцев. И у еще более далеких от животных амебоидных Holozoa они тоже встречаются. Нельзя же предполагать, что вообще у всех Holozoa когда-то была нервная система (то есть предположить-то можно, но это будет очевидный пример сведения к абсурду). Вот и пришлось переименовать синаптические белки в протосинаптические: ясно ведь, что у одноклеточных организмов их функции — иные, чем у обладателей нервных систем. Хотя сами белки — те же самые, многие из них сохраняются на всем протяжении эволюционного древа от одноклеточных амеб до позвоночных.
И у еще более далеких от животных амебоидных Holozoa они тоже встречаются. Нельзя же предполагать, что вообще у всех Holozoa когда-то была нервная система (то есть предположить-то можно, но это будет очевидный пример сведения к абсурду). Вот и пришлось переименовать синаптические белки в протосинаптические: ясно ведь, что у одноклеточных организмов их функции — иные, чем у обладателей нервных систем. Хотя сами белки — те же самые, многие из них сохраняются на всем протяжении эволюционного древа от одноклеточных амеб до позвоночных.
Что могут делать протосинаптические белки у одноклеточных организмов? Например, помогать концентрировать ионные каналы в определенных областях клеточной мембраны (белки GKAP), регулировать сборку микрофиламентов в клеточных выростах (белки Shank) или обеспечивать слияние движущихся внутри клетки мембранных пузырьков (белки SNARE). А уже после возникновения многоклеточности все эти функции белков пригодились для создания архитектуры синапсов. Следует добавить, что для многих протосинаптических белков (которые точно есть и у животных с нервной системой, и у одноклеточных Holozoa) исходные функции просто еще неизвестны — возможно, эти детали прояснятся в ближайшее время.
Таким образом, наличие нервной системы у предков губок и пластинчатых ничем серьезно не подтверждается. Что касается гребневиков, то они отличаются от стрекающих с билатериями и по набору нейромедиаторных систем, и по молекулярной архитектуре синапсов (она «собрана» на основе отличающегося набора белков), поэтому гипотеза о самостоятельном возникновении их нервной системы выглядит хорошо обоснованной независимо от положения гребневиков на филогенетическом древе. Смена гипотезы «первичности гребневиков» на гипотезу «первичности губок» в этом отношении ничего принципиально не меняет. Разбираться нужно именно с молекулярными компонентами нервных клеток, во всех деталях, чтобы в качестве промежуточного итога можно было реконструировать состояние, свойственное общему предку гребневиков, стрекающих и билатерий, и посмотреть — насколько сильно оно отличается от состояния губок. Тогда и можно будет попытаться окончательно выяснить, сколько раз происходил эволюционный прорыв на уровень «настоящих многоклеточных животных» (с нервной системой). На данный момент гипотеза, что он произошел дважды, остается достаточно вероятной.
На данный момент гипотеза, что он произошел дважды, остается достаточно вероятной.
Общим предком гребневиков, пластинчатых, стрекающих и билатерий (то есть всех животных, кроме губок) Нильсен считает уже знакомую нам геккелевскую гастрею — голопелагический организм, потомки которого довольно быстро в эволюционном масштабе времени (но независимо в двух разных линиях) приобрели нервную систему и мышцы. Гребневики так до сих пор и остались голопелагическими: у подавляющего большинства из них весь жизненный цикл проходит в плавающем состоянии. А вот предки стрекающих и билатерий в некоторый момент опустились на дно, сохранив, однако, плавающую личинку. Свойственная многим стрекающим жизненная форма медузы, а также многочисленные активно плавающие билатерии вплоть до морских стрелок, рыб и китов — это уже результаты вторичного возвращения в толщу воды.
Совершенно особый статус имеет ветвь пластинчатых. Ее положение на эволюционном древе до сих пор окончательно не установлено: на большинстве полученных деревьев пластинчатые выглядят сестринской группой по отношению к ветви, объединяющий стрекающих и билатерий, но есть и такие реконструкции, где пластинчатые находятся ближе к стрекающим, чем к билатериям (C. E. Laumer et al., 2018. Support for a clade of Placozoa and Cnidaria in genes with minimal compositional bias). Насколько велико значение этих «разночтений» — пока неясно. У пластинчатых нет ни рта, ни мышц, ни нервных клеток, но есть уникальная система межклеточной сигнализации, основанная на использовании разнообразных пептидов, то есть коротких цепочек аминокислот (F. Varoqueaux et al., 2018. High cell diversity and complex peptidergic signaling underlie placozoan behavior). Не исключено, что родственниками пластинчатых — только гораздо более крупными — были некоторые представители загадочной вендской фауны, родство которых с многоклеточными животными сейчас считается доказанным (см. Подтверждена принадлежность дикинсонии к животному царству, «Элементы», 24.09.2018). Нильсен считает пластинчатых потомками гастреи, которая легла на дно и распласталась по нему. При этом слой клеток, выстилавший первичный кишечник (архентерон), обратился в подошву, на которой животное теперь ползает. Тут можно только добавить, что очень похожие взгляды на природу пластинчатых высказывались и раньше (см.
E. Laumer et al., 2018. Support for a clade of Placozoa and Cnidaria in genes with minimal compositional bias). Насколько велико значение этих «разночтений» — пока неясно. У пластинчатых нет ни рта, ни мышц, ни нервных клеток, но есть уникальная система межклеточной сигнализации, основанная на использовании разнообразных пептидов, то есть коротких цепочек аминокислот (F. Varoqueaux et al., 2018. High cell diversity and complex peptidergic signaling underlie placozoan behavior). Не исключено, что родственниками пластинчатых — только гораздо более крупными — были некоторые представители загадочной вендской фауны, родство которых с многоклеточными животными сейчас считается доказанным (см. Подтверждена принадлежность дикинсонии к животному царству, «Элементы», 24.09.2018). Нильсен считает пластинчатых потомками гастреи, которая легла на дно и распласталась по нему. При этом слой клеток, выстилавший первичный кишечник (архентерон), обратился в подошву, на которой животное теперь ползает. Тут можно только добавить, что очень похожие взгляды на природу пластинчатых высказывались и раньше (см. В. В. Алешин, Н. Б. Петров, 2001. Регресс в эволюции многоклеточных животных).
В. В. Алешин, Н. Б. Петров, 2001. Регресс в эволюции многоклеточных животных).
В последней части своей статьи Нильсен касается проблемы соотношения результатов молекулярной филогенетики (основанных исключительно на чтении нуклеотидных текстов) с данными обычной морфологии, изучающей структуру организмов. Из двух обсуждаемых конкурирующих гипотез одна, а именно гипотеза «первичности гребневиков», не имеет морфологической поддержки: у гипотетической эволюционной ветви, включающей всех многоклеточных животных, кроме гребневиков, нет ни одного общего уникального (синапоморфного) структурного признака. Напротив, у гипотезы «первичности губок» морфологическая поддержка есть: тут можно без труда назвать уникальные признаки для каждой из ветвей, возникающих на эволюционном древе. Нильсен — ученый, глубоко преданный классической морфологии, и его явно радует тот факт, что гипотеза «первичности губок» сейчас побеждает.
Источник: Claus Nielsen. Early animal evolution: a morphologist’s view // Royal Society open science./i.s3.glbimg.com/v1/AUTH_59edd422c0c84a879bd37670ae4f538a/internal_photos/bs/2021/D/v/USroPzSIulsUgcBMGAFQ/e7uzqvovcamq3mk.jpg) 2019. V. 6. № 7. 190638.
2019. V. 6. № 7. 190638.
Сергей Ястребов
Ученые ДВФУ испытали природное соединение из морской губки против рака
Контактный центр
RU
EN
Версия для слабовидящих
9 марта 2021 — Новости ДВФУ
#Наука
Ученые Дальневосточного федерального университета (ДВФУ) вместе с российскими и немецкими коллегами продолжают цикл исследований противоопухолевых соединений, синтезированных на основе биологически активных молекул из морской губки Fascaplysinopsis reticulata. В статье, опубликованной в Marine Drugs, исследователи рассказали об активности одного из этих перспективных веществ против клеток рака, устойчивых к стандартной химиотерапии, а также объяснили особенности его механизма действия.
Ученые испытали биологическое действие алкалоида 3,10-дибромфаскаплизин на различных клетках рака простаты, в том числе устойчивых к химиотерапии стандартным препаратом. Это соединение впервые было выделено из морской губки Fascaplysinopsis reticulata и впоследствии химически синтезировано сотрудниками ДВФУ. Под действием вещества опухолевые клетки погибали по программируемому механизму. Это процесс называется апоптозом и считается наиболее благоприятным сценарием действия противоопухолевых лекарственных соединений.
«Интересно, что испытанное соединение, способствуя гибели раковых клеток, в том числе устойчивых к стандартной химиотерапии, одновременно активирует в них фермент, который опухолевые клетки защищает. В принципе, это нельзя оценивать в категориях «хорошо» или «плохо». Это просто механизм действия, понимая который, 3, 10-дибромфаскаплизин нужно применять вместе с молекулами, подавляющими активность этих ферментов», — рассказал сотрудник лаборатории биологически активных веществ Школы естественных наук ДВФУ, старший научный сотрудник лаборатории фармакологии Национального научного центра морской биологии Дальневосточного отделения Российской академии наук Сергей Дышловой.
Ученый рассказал, что помимо самостоятельной противоопухолевой активности, синтезированное соединение в сочетании с уже известными химиотерапевтическими препаратами усиливает их действие.
На следующем этапе планируется проверить действие 3,10–дибромфаскаплизина на неопухолевых клетках. Работы в этом направлении ведутся и уже поддержаны Российским фондом фундаментальных исследований, ученые планируют опубликовать результаты в течение 2021 года.
«Фаскаплизины достаточно токсичны для здоровых клеток. В нашей лаборатории мы пытаемся так модифицировать структуру этих соединений, чтобы уменьшить их цитотоксическое воздействие на обычные клетки, сохранив при этом необходимый противоопухолевый эффект. Цель — создать вещество для таргетированной терапии, с минимумом побочных эффектов для здоровых клеток организма», — отметил заведующий кафедрой органической химии Школы естественных наук ДВФУ Максим Жидков.
По словам ученых, о создании лекарства можно говорить на горизонте 10-15 лет, учитывая необходимость длительных предварительных и клинических испытаний.
В исследовании приняли участие специалисты ДВФУ, Национального научного центра морской биологии, Мартини Клиник, Университетской клиники Гамбург-Эппендорф (Германия), НИИ биомедицинской химии имени В.Н. Ореховича, Института Молекулярной Биологии им. В.А. Энгельгардта РАН.
Разработка инновационных и перспективных методов борьбы с заболеваниями является одним из приоритетных научных направлений для Дальневосточного федерального университета. Особое внимание уделяется исследованию современных методов диагностики и лечения рака и других трудноизлечимых болезней на молекулярном и генетическом уровнях. Работа ведется на собственной научной базе университета и в сотрудничестве с крупнейшими научно-исследовательскими центрами мира.
Источником новых потенциальных лекарственных соединений инновационных соединений является Мировой океан, изучение которого — одно из приоритетных направлений научно-исследовательской деятельности ДВФУ, развиваемое в тесном сотрудничестве с Российской академией наук, отечественными и зарубежными партнерами.
Пресс-служба ДВФУ,
[email protected]
26.09.2022
День первокурсника
21.09.2022
День первокурсника
16 сентября
#Наука
Исследование преподавателя ДВФУ по истории религиозных общин войдет в серию Studia Religiosa
15 сентября
#Наука
Перспективы программы Тихоокеанского Плавучего университета обсудили в ДВФУ
12 сентября
#Наука
Ученые ДВФУ начали новый этап по исследованию экосистем Камчатки
Ученые превратили морскую губку в графит
https://ria.ru/20191017/1559897092. html
html
Ученые превратили морскую губку в графит
Ученые превратили морскую губку в графит — РИА Новости, 17.10.2019
Ученые превратили морскую губку в графит
Уникальное свойство морской губки не разрушаться при нагревании, а переходить в графит обнаружилось в результате эксперимента, проведенного российскими и… РИА Новости, 17.10.2019
2019-10-17T14:42
2019-10-17T14:42
2019-10-17T14:42
наука
сыктывкарский государственный университет
санкт-петербургский государственный университет
санкт-петербургский университет информационных технологий
открытия — риа наука
/html/head/meta[@name=’og:title’]/@content
/html/head/meta[@name=’og:description’]/@content
https://cdnn21.img.ria.ru/images/155989/58/1559895810_0:229:1229:920_1920x0_80_0_0_1272a23e44e84dec11164343dfeb3525.png
МОСКВА, 17 окт — РИА Новости. Уникальное свойство морской губки не разрушаться при нагревании, а переходить в графит обнаружилось в результате эксперимента, проведенного российскими и зарубежными учеными. Это открывает путь к получению новых экологически чистых материалов, сохраняющих прочность при очень высоких температурах. Результаты исследования опубликованы в журнале Science Advances.При разработке новых материалов ученые часто черпают идеи у природы. В бионике — науке о применении функций и структур организмов в технических устройствах — есть отдельное направление, изучающее возможности живых существ, приспосабливаться к экстремальным условиям. Один из объектов экстремальной бионики — морские губки, которые за 600 миллионов лет своей истории сумели пережить самые разнообразные природные катаклизмы. Секрет невероятной стойкости морских губок — в веществе, из которого состоит их скелет, — спонгине.Спонгин устойчив к воздействию кислот, а также выдерживает нагревание до 360 °C при наличии кислорода и до 1200 °C в бескислородной среде. Это делает его перспективным сырьем для создания композитных материалов. Спонгин состоит из рогового вещества. Из него же состоят волосы, рога и когти.
Это открывает путь к получению новых экологически чистых материалов, сохраняющих прочность при очень высоких температурах. Результаты исследования опубликованы в журнале Science Advances.При разработке новых материалов ученые часто черпают идеи у природы. В бионике — науке о применении функций и структур организмов в технических устройствах — есть отдельное направление, изучающее возможности живых существ, приспосабливаться к экстремальным условиям. Один из объектов экстремальной бионики — морские губки, которые за 600 миллионов лет своей истории сумели пережить самые разнообразные природные катаклизмы. Секрет невероятной стойкости морских губок — в веществе, из которого состоит их скелет, — спонгине.Спонгин устойчив к воздействию кислот, а также выдерживает нагревание до 360 °C при наличии кислорода и до 1200 °C в бескислородной среде. Это делает его перспективным сырьем для создания композитных материалов. Спонгин состоит из рогового вещества. Из него же состоят волосы, рога и когти. Его особые свойства связаны не столько с составом, сколько с необычной внутренней структурой. Волокна рогового вещества толщиной до 100 мкм объединены в нем в сложные трехмерные системы с высокой пористостью. Применение спонгина открывает широкие возможности для создания возобновляемых, нетоксичных и биоразлагаемых органических каркасов сантиметрового и даже метрового размера. Современные технологии позволяют создавать такие структуры только на наноуровне.Для повышения прочности органических каркасов перед промышленным использованием их подвергают карбонизации — высокотемпературному насыщению углекислым газом. Чтобы понять, как поведут себя спонгиновые структуры в таких условиях, ученые решили изучить свойства скелетов морских губок при нагревании.Работа проводилась международной командой исследователей, в которую входили ученые из Фрайбергской горной академии (Германия), университетов США, Польши, Франции, Германии, Словакии. С российской стороны участвовали ученые из Санкт-Петербургского государственного университета, Сыктывкарского государственного университета и Университета ИТМО (Санкт-Петербург).
Его особые свойства связаны не столько с составом, сколько с необычной внутренней структурой. Волокна рогового вещества толщиной до 100 мкм объединены в нем в сложные трехмерные системы с высокой пористостью. Применение спонгина открывает широкие возможности для создания возобновляемых, нетоксичных и биоразлагаемых органических каркасов сантиметрового и даже метрового размера. Современные технологии позволяют создавать такие структуры только на наноуровне.Для повышения прочности органических каркасов перед промышленным использованием их подвергают карбонизации — высокотемпературному насыщению углекислым газом. Чтобы понять, как поведут себя спонгиновые структуры в таких условиях, ученые решили изучить свойства скелетов морских губок при нагревании.Работа проводилась международной командой исследователей, в которую входили ученые из Фрайбергской горной академии (Германия), университетов США, Польши, Франции, Германии, Словакии. С российской стороны участвовали ученые из Санкт-Петербургского государственного университета, Сыктывкарского государственного университета и Университета ИТМО (Санкт-Петербург). Исследование поддержано Российским научным фондом.Ученые провели эксперимент по нагреванию скелета морской губки до 1200 градусов. Другие биологические материалы при такой температуре превращаются в бесполезный угольный порошок. Но скелет губки после нагревания в течение часа полностью сохранил свою микро- и наноструктуру, хотя и потерял 70% объема. Изучение с помощью электронного микроскопа показало, что спонгин полностью превратился в графитовую губку, то есть сохранил свой внешний вид, но внутри заместился графитом. При этом он остался настолько прочным, что из него удалось вырезать разнообразные фигуры.
Исследование поддержано Российским научным фондом.Ученые провели эксперимент по нагреванию скелета морской губки до 1200 градусов. Другие биологические материалы при такой температуре превращаются в бесполезный угольный порошок. Но скелет губки после нагревания в течение часа полностью сохранил свою микро- и наноструктуру, хотя и потерял 70% объема. Изучение с помощью электронного микроскопа показало, что спонгин полностью превратился в графитовую губку, то есть сохранил свой внешний вид, но внутри заместился графитом. При этом он остался настолько прочным, что из него удалось вырезать разнообразные фигуры.
https://ria.ru/20190722/1556759576.html
https://ria.ru/20170707/1498047749.html
РИА Новости
1
5
4.7
96
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
2019
РИА Новости
1
5
4.7
96
internet-group@rian. ru
ru
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
Новости
ru-RU
https://ria.ru/docs/about/copyright.html
https://xn--c1acbl2abdlkab1og.xn--p1ai/
РИА Новости
1
5
4.7
96
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
1920
1080
true
1920
1440
true
https://cdnn21.img.ria.ru/images/155989/58/1559895810_0:48:1229:970_1920x0_80_0_0_6f5d1e42548553736fdfc5d63a6d04d1.png
1920
1920
true
РИА Новости
1
5
4.7
96
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
РИА Новости
1
5
4.7
96
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og. xn--p1ai/awards/
xn--p1ai/awards/
сыктывкарский государственный университет, санкт-петербургский государственный университет, санкт-петербургский университет информационных технологий, открытия — риа наука
Наука, Сыктывкарский государственный университет, Санкт-Петербургский государственный университет, Санкт-Петербургский университет информационных технологий, Открытия — РИА Наука
МОСКВА, 17 окт — РИА Новости. Уникальное свойство морской губки не разрушаться при нагревании, а переходить в графит обнаружилось в результате эксперимента, проведенного российскими и зарубежными учеными. Это открывает путь к получению новых экологически чистых материалов, сохраняющих прочность при очень высоких температурах. Результаты исследования опубликованы в журнале Science Advances.
При разработке новых материалов ученые часто черпают идеи у природы. В бионике — науке о применении функций и структур организмов в технических устройствах — есть отдельное направление, изучающее возможности живых существ, приспосабливаться к экстремальным условиям.
Один из объектов экстремальной бионики — морские губки, которые за 600 миллионов лет своей истории сумели пережить самые разнообразные природные катаклизмы. Секрет невероятной стойкости морских губок — в веществе, из которого состоит их скелет, — спонгине.
Спонгин устойчив к воздействию кислот, а также выдерживает нагревание до 360 °C при наличии кислорода и до 1200 °C в бескислородной среде. Это делает его перспективным сырьем для создания композитных материалов.
22 июля 2019, 12:28Наука
Российские физики открыли новое необычное свойство графена
Спонгин состоит из рогового вещества. Из него же состоят волосы, рога и когти. Его особые свойства связаны не столько с составом, сколько с необычной внутренней структурой. Волокна рогового вещества толщиной до 100 мкм объединены в нем в сложные трехмерные системы с высокой пористостью.
Применение спонгина открывает широкие возможности для создания возобновляемых, нетоксичных и биоразлагаемых органических каркасов сантиметрового и даже метрового размера. Современные технологии позволяют создавать такие структуры только на наноуровне.
Современные технологии позволяют создавать такие структуры только на наноуровне.
Для повышения прочности органических каркасов перед промышленным использованием их подвергают карбонизации — высокотемпературному насыщению углекислым газом. Чтобы понять, как поведут себя спонгиновые структуры в таких условиях, ученые решили изучить свойства скелетов морских губок при нагревании.
7 июля 2017, 15:25Наука
Биологи нашли лекарство от «спящего» туберкулеза в морских губках
Работа проводилась международной командой исследователей, в которую входили ученые из Фрайбергской горной академии (Германия), университетов США, Польши, Франции, Германии, Словакии. С российской стороны участвовали ученые из Санкт-Петербургского государственного университета, Сыктывкарского государственного университета и Университета ИТМО (Санкт-Петербург). Исследование поддержано Российским научным фондом.
Ученые провели эксперимент по нагреванию скелета морской губки до 1200 градусов. Другие биологические материалы при такой температуре превращаются в бесполезный угольный порошок. Но скелет губки после нагревания в течение часа полностью сохранил свою микро- и наноструктуру, хотя и потерял 70% объема. Изучение с помощью электронного микроскопа показало, что спонгин полностью превратился в графитовую губку, то есть сохранил свой внешний вид, но внутри заместился графитом. При этом он остался настолько прочным, что из него удалось вырезать разнообразные фигуры.
Другие биологические материалы при такой температуре превращаются в бесполезный угольный порошок. Но скелет губки после нагревания в течение часа полностью сохранил свою микро- и наноструктуру, хотя и потерял 70% объема. Изучение с помощью электронного микроскопа показало, что спонгин полностью превратился в графитовую губку, то есть сохранил свой внешний вид, но внутри заместился графитом. При этом он остался настолько прочным, что из него удалось вырезать разнообразные фигуры.
© Фото : Iaroslav Petrenko et al. / Science Advances, 2019.Графитизированный скелет губки в разрезе
© Фото : Iaroslav Petrenko et al. / Science Advances, 2019.
Графитизированный скелет губки в разрезе
«Сейчас, когда проблема загрязнения окружающей среды стала одной из главных для всей планеты, появление новых материалов крайне важно. Использовать спонгин экономически целесообразно, так как это естественный, возобновляемый и экологически чистый ресурс, который можно производить по всему миру. Уже сейчас в странах Средиземноморья, Австралии и США выращивают кератозные губки. При правильном управлении развитие этих технологий не повредит популяции морских животных, а материалы из спонгина можно будет применять в самых разных областях промышленности», — приводит пресс-служба РНФ слова Александра Ересковского, профессора кафедры эмбриологии биологического факультета Санкт-Петербургского государственного университета.
Уже сейчас в странах Средиземноморья, Австралии и США выращивают кератозные губки. При правильном управлении развитие этих технологий не повредит популяции морских животных, а материалы из спонгина можно будет применять в самых разных областях промышленности», — приводит пресс-служба РНФ слова Александра Ересковского, профессора кафедры эмбриологии биологического факультета Санкт-Петербургского государственного университета.
|
|
 Без них не
Без них не Да и
Да и Никакой реакции со стороны губки не
Никакой реакции со стороны губки не
 4).
4). Кроме того, с током воды мимо хоаноцитов проплывают мелкие (не более
Кроме того, с током воды мимо хоаноцитов проплывают мелкие (не более Любопытно, что
Любопытно, что Губки – гермафродиты. Половые клетки у них образуются
Губки – гермафродиты. Половые клетки у них образуются У некоторых губок дело происходит
У некоторых губок дело происходит
 И все же, видимо, губок стоит считать многоклеточными животными. Очень
И все же, видимо, губок стоит считать многоклеточными животными. Очень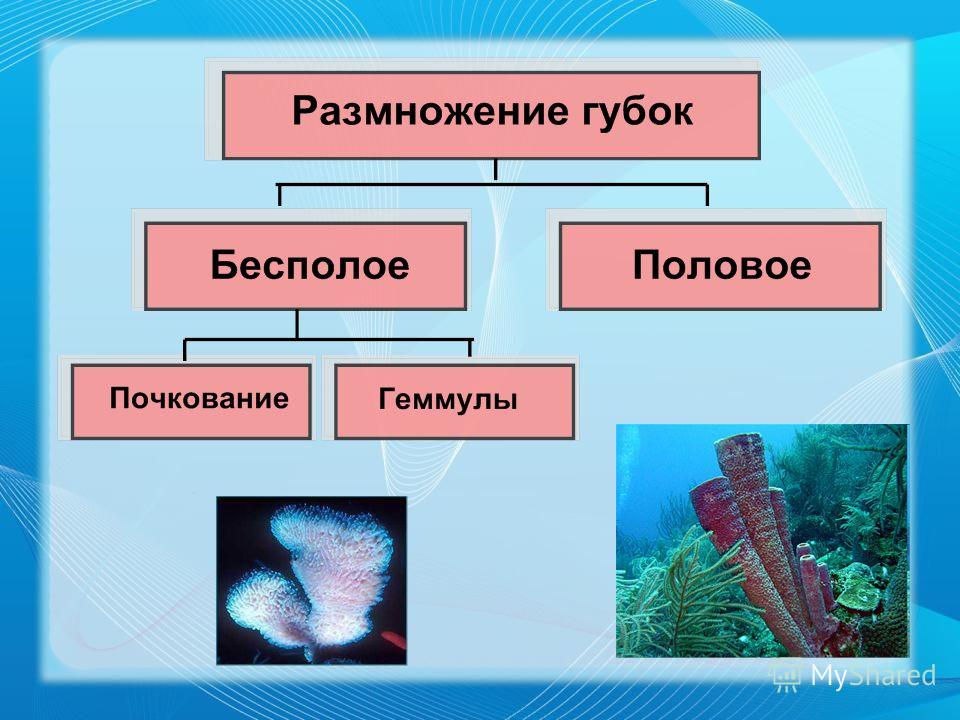
 8). У тех губок, у которых скелет представлен только
8). У тех губок, у которых скелет представлен только Многие губки, содержащие йод, помогают при лечении базедовой
Многие губки, содержащие йод, помогают при лечении базедовой Ученик и ассистент великого Эрнста Геккеля, он много занимался
Ученик и ассистент великого Эрнста Геккеля, он много занимался И. Мечникова, дальше
И. Мечникова, дальше
Чих чихом, губки наполняют моря своей слизью
Наука|Чихание чихом, губки наполняют моря своей слизью
https://www.nytimes.com/2022/08/10/science/sea-sponges -sneezing.html
Реклама
Продолжить чтение основной истории
Трилобиты
У вас может возникнуть соблазн сказать «gesundheit», но сопли морских существ помогают питаться другим морским организмам.
Видео
Чихающая индо-тихоокеанская губка Chelonaplysilla. Видео от Current Biology/Kornder et al.
Автор Сэм Джонс
Чихание далеко не исключительно человеческое поведение. Возможно, вы видели, как это делает ваша собака или кошка, или смотрели видео на YouTube, где жираф чихает на ничего не подозревающего малыша в зоопарке. Фактически, чихание даже не требует нервной системы, не говоря уже о носе, и восходит к некоторым из первых многоклеточных животных: губкам.
Губка существует не менее 600 миллионов лет. «Это самое успешное животное, о котором я знаю, потому что оно такое старое и повсюду», — сказал Джаспер де Гой, морской эколог из Амстердамского университета. Как фильтраторы, губки играют решающую роль в своих водных экосистемах, всасывая воду, наполненную разнообразными органическими веществами, перерабатывая ее и выпуская в виде отходов, которыми питаются такие организмы, как улитки, офиуры и трубчатые черви. «Губка — это, по сути, животное, у которого много маленьких ртов и одно или несколько больших выходных отверстий», — сказал доктор де Гой. Эти «устья» называются ostia, а отверстия, через которые вытекает вода, — oscula.
«Губка — это, по сути, животное, у которого много маленьких ртов и одно или несколько больших выходных отверстий», — сказал доктор де Гой. Эти «устья» называются ostia, а отверстия, через которые вытекает вода, — oscula.
В течение многих лет ученые знали, что губки могут регулировать поток воды с помощью многоминутного сокращения тела — т. е. «чихания», — но теперь доктор де Гой и его коллеги обнаружили, что губки чихают форма самоочищения, высвобождающая частицы отходов в слизи через их отверстия. Работа была опубликована в Current Biology в среду.
Исследователи столкнулись с губками, чихающими соплями, во время работы над проектом по изучению роли губок в перемещении питательных веществ через экосистему рифа. Работа потребовала, чтобы Никлас Корндер, другой морской эколог из Амстердама, провел много времени с губками. «Я проводил целые дни, просто глядя на их поверхность; это было довольно скучно», — вспоминал он. (В то время г-н Корндер занимался подводным плаванием в Карибском море. )
)
Изображение
Все три вида морских губок, вероятно, чихают прямо сейчас. Кредит… Бенджамин Мюллер
К счастью, все стало интереснее, когда он начал видеть непрозрачный волокнистый материал, исходящий от губок. «Тогда я возвращался к этому позже, и волокнистые вещи исчезали», — сказал он.
Чтобы выяснить, что это за «волокнистые штуки», исследователи записали покадровую съемку губок, в частности карибской трубчатой губки Aplysina archeri. В лаборатории они смогли идентифицировать нити как потоки слизи, несущие отходы. Они выходили из устьев губки, двигались по поверхности организма и собирались в комки, которые можно было высвободить при чихании, а затем быстро поглощали другие морские твари.
При первом просмотре замедленной съемки Юки Эссер — в то время аспирантка факультета биоинформатики в Амстердаме и соавтор исследования — была разочарована, думая, что движение, которое она видела (например, чих), было просто камерой. ошибка фокусировки. «Я подумала, что причиной этого должна быть капля воды или что-то еще на объективе камеры», — сказала она. Но вскоре она поняла, что это не ошибка. И как только г-жа Эссер и ее коллеги обнаружили, что они сняли почти идентичное покадровое видео A. archeri у побережья Кюрасао, видеосъемка «стала чем-то вроде спорта», — сказала она. «Типа: «Может быть, мы засняли еще один чих на камеру!»
ошибка фокусировки. «Я подумала, что причиной этого должна быть капля воды или что-то еще на объективе камеры», — сказала она. Но вскоре она поняла, что это не ошибка. И как только г-жа Эссер и ее коллеги обнаружили, что они сняли почти идентичное покадровое видео A. archeri у побережья Кюрасао, видеосъемка «стала чем-то вроде спорта», — сказала она. «Типа: «Может быть, мы засняли еще один чих на камеру!»
Видео
Замедленные и замедленные кадры чихания трубчатой губки Аплисины. Видео Kornder et al., Current Biology
Исследователи считают, что чихание слизи, содержащей отходы, является широко распространенной тактикой среди губок во всем мире. И исследование вызывает больше вопросов, говорит Салли Лейс, биолог-эволюционист из Университета Альберты и соавтор исследования.
— Слизь, — сказала она. «Похожа ли она на слизь других животных? И какие клетки его производят? Она также хочет знать, что вызывает чихание. «Когда у нас течет из носа, мы достаем салфетки», — сказала она. «Но откуда губка знает, что сейчас самое время чихнуть?»
«Но откуда губка знает, что сейчас самое время чихнуть?»
Изучение этой слизи может улучшить понимание учеными того, как микробы и, возможно, болезни передаются в рифовых экосистемах, сказал Блейк Усидзима, изучающий кораллы в Университете Северной Каролины в Уилмингтоне и не участвовавший в новом исследовании. Он также поражен тем, что это исследование может рассказать нам о нашей собственной эволюции.
«Это может дать нам представление о том, как ранняя жизнь превратилась из этих мягких безмозглых существ в эти сложные организмы, строящие космические корабли», — сказал доктор Ушиджима.
Sponges: The Original Animal House
Итак, вы — группа сестринских клеток, стремящихся собраться вместе и создать первый в мире кооператив для животных, место, где вы и ваши приятели сможете жить вместе в маленьком социалистическом утопия и специализируются на выполнении одной работы, а не пытаются делать все сразу, как эти глупые, одноклеточные, суровые индивидуалисты-протисты. Как это может выглядеть?
Как это может выглядеть?
Так уж получилось, что он, вероятно, был очень похож на губку, потому что они или что-то очень похожее на них, вероятно, были первыми. А вот как они выглядят сегодня:
Губка в виде трубки, губка для вазы, губка из веревки и губка для корочки входят в бар. Подожди, поцарапай часть бара. Губки бывают разных форм! Губка трубчатая желтая, Aplysina fistularis, губка пурпурная, Niphates digitalis, корковая губка красная, Spiratrella coccinea, и губка серая, Callyspongia sp. в Карибском море, недалеко от Каймановых островов. 23 мая 2007 г. Экспедиционная группа Creative Commons Twilight Zone 2007 г., NOAA-OE. Нажмите для ссылки.
Какое-то позвоночное животное прячется внутри великолепной синей бочкообразной губки у побережья Бонайре в Карибском море. Creative Commons Крис Коккаро, NOAA-OE. Щелкните изображение для ссылки.
Губка — это, по существу, многоклеточный организм без органов и тканей, но с специализированными клетками, что отличает ее от мелких многоклеточных протистов. Как и у водорослей, тонкость строения тела губки и тот факт, что большая часть ее клеток подвергается воздействию циркулирующей воды (поставщик кислорода и пищи, удаляющая отходы), делает ненужными такие органы, как сердце, почки и пищеварительный тракт.
Типичная бочкообразная или трубчатая губка, по сути, представляет собой перфорированный мешок. Нажмите здесь, чтобы увидеть красивую диаграмму, которая поможет проиллюстрировать следующий текст. Снаружи губки покрыты эпидермальными клетками; в стенке губки находятся блуждающие амебоидные клетки, которые выполняют некоторые обязанности, подобные иммунной системе, и выделяют кристаллические спикулы (обычно карбонат кальция или кремневую кислоту) или белковые волокна, поддерживающие губку; специальные клетки, называемые пороцитами, могут окружать впадающие в стенку поры; и характерные хоаноциты или клетки с воротничком выстилают внутренние пространства губки. Спикулы могут иметь любое количество точек; окаменелости стеклянной губки Hydnoceras из девона (390-360 млн лет назад) выглядят во всем мире так, как будто они завернуты в плед благодаря своим шестиконечным спикулам, которые кажутся четырехконечными в двух измерениях, видимых на его внешней стороне.
Спикулы губки. Некоторые могут быть четырех- или шестиконечными; другие зацепились. Общественное достояние, нажмите для ссылки.
Воротнички, давшие название хоаноцитам, состоят из пальцевидных отростков цитоплазматической мембраны, называемых микроворсинками (похожих на те, что выстилают тонкую кишку), окружающих хвостовидные жгутики. Бьющиеся жгутики создают ток, который втягивает воду через пороциты или межклеточные поры (в любом случае отверстия называются «устьями») и наружу через центральную полость губки, называемую оскулумом. Частицы пищи, которые натыкаются на микроворсинки, прилипают к ним и либо поглощаются и перевариваются, либо передаются амебоидным клеткам, которые выполняют свою работу.
Вы можете видеть, что хоаноциты легче всего генерируют поток воды, когда нерестятся губки. В этом видео водолазы снимают нерест губок в Национальном морском заповеднике Флауэр Гарден Бэнкс в Мексиканском заливе, примерно в 70–115 милях от побережья Техаса и Луизианы.
Губки в конце видео несомненно выделяют сперму. Первая группа губок может состоять из самок, откладывающих яйца, но они, скорее всего, являются гермафродитами (большинство губок) и откладывают оплодотворенные яйца или личинки. Сперматозоиды, образованные в результате трансмогрификации хоаноцитов, должны найти другую губку соответствующего вида и слиться с хоаноцитом внутри. Вот где это становится триповым. Оказавшись там, хоаноцит, несущий сперму, превращается в амебу и ползет к яйцеклетке. Сама яйцеклетка образовалась, когда либо хоаноцит, либо амебоцит поглотил специальные «кормящие» клетки, образовав желток. Амебоидный хоаноцит, несущий сперму, и набухшее яйцо сливаются. Как правильные хромосомы находят друг друга, чтобы сформировать одно диплоидное ядро во всем этом беспорядке, не заканчивая пятью или шестью копиями генома, это тема для другого дня.
Затем губка может выпустить оплодотворенные яйца или сохранить их. В любом случае, зигота превращается в личинку, состоящую из шара клеток с внешними жгутиками или ресничками, которые управляют всем этим. Они какое-то время возятся, молясь, чтобы их не съели, пока ищут подходящее место для постройки своей хоанофлагеллятной квартирки, и вуаля, личинка приземляется и превращается в губку.
Губки часто считаются колониальными организмами, такими как водоросли Volvox . Клетки настолько слабо связаны, что если вы пропустите губку через фильтр и перемешаете клетки, они легко снова ассоциируются, образуя другую губку. Слизевики, хотя и эволюционировали из совершенно другой амебоидной линии, тоже могут проделывать этот трюк (попробуйте *это* с любым предположительно «высшим» животным).
Это подводит нас к сути этого сообщения в блоге и к причине, по которой я включил его в сегодняшний блог-марафон «Друзья в группах». Предки сегодняшних губок, возможно, были первыми по-настоящему многоклеточными животными. В С.Е. Блог Лабораторной крысы Гулда — это хорошее определение многоклеточности, в котором говорится, что клетки многоклеточных организмов должны держаться вместе, общаться друг с другом, зависеть друг от друга в своем выживании и специализироваться. Губки, похоже, соответствуют всем этим критериям. Справедливости ради следует отметить, что placazoans, очаровательно простая бродячая клеточная пленка, также претендуют на эту честь, но я должен отложить их обсуждение на другой день.
Вопрос о том, как давно это произошло, является предметом дискуссий, но кажется, что это было по крайней мере 543 миллиона лет назад, во время протерозоя, откуда мы знаем о первых ископаемых спикулах губок, а может быть и намного больше. Ранние губки, возможно, не имели спикул или каких-либо других следов, которые могли бы окаменеть. Всякий раз, когда они появлялись, они, должно быть, значительно очищали то, что, вероятно, становилось довольно густой морской водой, поскольку они были и, по сути, являются маленькими установками фильтрации океана, первыми из много питателей-фильтров.
Независимо от того, кто был первым многоклеточным животным, губки и плакозои, вероятно, не были первыми животными . Это различие может принадлежать протистам, называемым хоанофлагеллятами, которые представляют собой одноклеточные или многоклеточные букеты или сферы того, что выглядит точно так же, как хоаноциты внутри губок. По крайней мере, одна свободноживущая сферическая колония хоанофлагеллят (по совпадению названная Sphaeroeca volvox ) мало чем отличается от личинки губки.
Sphaeroeca sp., колониальная хоанофлагеллята. Общественное достояние, щелкните изображение для ссылки.
Еще у других видов хоанофлагеллят жгутиковые клетки обращены *внутрь* в полость, очень похожую на взрослую губку.
Но, может быть, вы спросите, откуда мы знаем, что такие малоподвижные существа вообще являются животными? Что ж, последовательности их ДНК говорят нам об этом сегодня. Но биологи знали, что губки являются животными гораздо дольше, чем мы знали о ДНК. Помните те белковые эластичные скелеты? (если вы когда-либо сжимали настоящую губку для ванны, вы знакомы с этим). Помимо прочего, эти волокна состоят из формы белка коллагена — того же белка в ваших сухожилиях, связках и коже, где, когда он ломается, появляются морщины. И хотя ни у одной другой группы животных нет настоящих хоаноцитов, у большинства основных групп (типов) есть «воротничковые клетки», и некоторые из них вполне могли произойти от предков хоаноцитов. И ваша иммунная система также содержит амебоидные «макрофаги», которые блуждают по вашему телу по своему желанию, проникая в ткани и выскальзывая из них, когда им заблагорассудится, в поисках захватчиков. Несомненно, есть и другие сходства.
Губки были первыми многоклеточными кооперативами животных, т.е. Некоторые из этих свободных клеточных федераций, подвергаясь сильному отбору и другим эволюционным силам, превращались в сплоченные многоклеточные кооперативные машины, такие как вы: остальные животные. Их клетки стали настолько связанными друг с другом, что мы больше не думаем о них как о клеточных кооперативах. О животных говорят как об одном «организме». Тем не менее, когда вы теряете клетки кожи или пищеварительные клетки, или когда какие-либо клетки в вашем теле добровольно умирают, не оставляя потомства, они приносят себя в жертву на благо этой более крупной, хотя и генетически идентичной сущности.
Прежде чем мы закончим с губками, давайте вкратце рассмотрим некоторые из наиболее забавных видов.
Одной из трех основных групп губок являются стеклянные губки, отличающиеся своими стеклянными шестиконечными кремнистыми спикулами и иногда изящным скульптурным внешним видом. Вот один из них, сохранившийся в зале Сант-Оушен в Смитсоновском институте. Обратите внимание на его длинные укореняющиеся спикулы, характерные для многих губок. Они были бы покрыты грязью в океане.
Цветочная корзина Венеры, стеклянная губка. В воде они тоже выглядят примерно так. Creative Commons Райан Сомма. Нажмите Изображение для ссылки.
Когда губки живут глубоко — очень глубоко — за пределами области, где кормление фильтром является прибыльным бизнесом, они, как ни удивительно, становятся плотоядными. Можно было бы не ожидать такого подвига от организма, который обычно имеет всю подвижность растения в горшке, но опять же, растения сами произвели немало плотоядных.
Одним из самых причудливых организмов на планете является плотоядная губка. Не верите мне? Вот губка для пинг-понга, Chondrocladia lampoglobalis, — обитатель морских глубин, напоминающий люстру в стиле ретро 60-х, популярную среди хипстеров-декораторов. Эти диковинки обычно имеют высоту около 50 см (20 дюймов) и обитают на глубине от 2600 до 3000 м (от 8500 до 9800 футов). Маленькие ракообразные или черви, которым не повезло приземлиться на древесную губку для пинг-понга, оказываются прикрепленными к ней крошечными спикулами, похожими на крюки для захвата. Это заставляет пищеварительные клетки мигрировать в это место и следующие несколько дней пережевывать пищу. Когда ужин закончен, они возвращаются на свои исходные места.
Другие плотоядные губки используют подобное вооружение. Помимо крючковатых спикул, они могут использовать липкие нити, чтобы запутывать и переваривать добычу. Большинство плотоядных губок потеряли свои хоаноциты и, как следствие, гидравлические системы, поскольку они были бы бессмысленны при отсутствии фильтрующего питания. Chondrocladia нет. Он использует сильно модифицированную гидравлическую систему для надувания шариков для пинг-понга на концах своих ответвлений.
Несколько видов губок — около 50 видов — живут в пресной воде. Мой друг Джордж Уотсон, технический дайвер, сделал прекрасные фотографии этой пресноводной губки глубоко — вероятно, за пределами досягаемости света — в маленьком, но *очень глубоком* озере в штате Нью-Мексико. Это, вероятно, вид Spongilla (и если кто-то знает точный вид, Джордж хотел бы знать), тот же род, что и этот экземпляр Spongilla lacustris , пойманный в верховьях реки Колумбия в штате Вашингтон:
Фотосинтезирующий, симбиотический пресноводный губка, Spongilla lacustris. Creative Commons Курт Л. Онтхинк. Щелкните изображение для ссылки.
Вы заметите, что он зеленый. Это потому, что у него, как и у многих пресноводных губок, есть симбионт зеленых водорослей. Некоторые морские виды сотрудничают с сине-зелеными водорослями (также называемыми цианобактериями) или даже с фотосинтезирующими динофлагеллятами (разновидность протистов), которые могут обеспечить более половины своих энергетических потребностей в местах, где питание фильтром не так уж и велико. Ааа, биология. Как я люблю тебя.
Наконец, я не могу закончить этот пост, не упомянув о чуме картофельных губок, выброшенных на берег после урагана Айрин в этом году. Бедные, невзрачные существа, кажется, были смыты штормом с морского дна, откуда их вынесло, умирая, на берег только для того, чтобы обвинить в том, что они зловонные и отвратительные. Если бы твой труп плавал несколько дней в Чесапикском заливе, держу пари, он бы тоже не так благоухал.
Выраженные взгляды принадлежат автору (авторам) и не обязательно совпадают с мнением Scientific American.
ОБ АВТОРАХ
Дженнифер Фрейзер, , научный писатель, лауреат премии AAAS в области научной журналистики, автор блога The Artful Amoeba для Scientific American . У нее есть ученые степени в области биологии, патологии растений и научного письма. Подпишитесь на Дженнифер Фрейзер в Твиттере. Автор: Ник Хиггинс
Факты о рыбе-губке | Demospongiae
Demospongiae
Последнее обновление: 8 мая 2022 г.
Проверено: AZ Animals Staff
V.Devolder/Shutterstock.com
Some people use sponges as sponges
Sponge Facts
- Main Prey
- Plankton, Mollusks, Crustaceans
- Group Behavior
- Colony
- Fun Fact
- Some people use sponges as sponges
- Расчетная численность популяции
- неизвестно
- Самая большая угроза
- Изменение климата, разрушение среды обитания
- Наиболее отличительная черта
- Поверхностные поры
- Habitat
- oceans, seas, lakes
Sponge Physical Characteristics
- Color
- Brown
- Yellow
- Red
- Blue
- Green
- Orange
- Skin Type
- Porous
- Срок службы
- 15–30 лет
- Вес
- 20 фунтов
- Длина
- 0,25–2 м (0,8–6 футов)
Покупка через них помогает нам продвигать миссию A-Z Animals по просвещению о видах в мире. .
Фотографии губок
Посмотрите все наши фотографии губок в галерее.
V.Devolder/Shutterstock.com
Посмотреть все изображения Губки!
Найдите своих любимых животных!
Губки — одни из самых простых и, возможно, древнейших животных на всей планете.
Это морские животные, которых легко спутать с растениями из-за отсутствия у них нервной системы, внутренних органов и подвижности. Все губки принадлежат к таксономическому типу Porifera, который является частью царства Animalia и включает более 500 родов и от 5000 до 10000 различных видов. Все губки являются водными животными, подавляющее большинство которых живет в соленой воде.
Невероятное животное: 4 факта о губке!
- Открытая циркуляция : В отличие от большинства животных, губки имеют открытую систему кровообращения, работа которой зависит от движения воды. Течения проталкивают воду через открытые поры и внутренние каналы, которые позволяют дышать, питаться и удалять отходы.

- Гибкое размножение : Губки ведут как половое, так и бесполое размножение. Многие из них являются гермафродитами, причем некоторые из них последовательно переключаются между мужскими и женскими ролями.
- Название, подходящее для : научное название типа губок, Porifera, буквально означает «носитель пор».
- Медленнодвижущиеся : Несмотря на то, что взрослые губки в основном являются неподвижными животными, они могут очень медленно передвигаться по поверхностям в процессе транспортировки клеток.
Классификация губок и научное название
Все губки относятся к типу Porifera, что в переводе с латыни означает «носители пор» или «носители пор». Это название происходит от множества видимых пор, покрывающих их поверхность. Этот тип делится на четыре класса: Calcarea Carbonica, Hexactinellida, Demospongiae и Homoscleromorpha. Общее название «губка» на самом деле восходит к древнегреческому.
Виды губок
Из-за множества уникальных характеристик по сравнению со всеми другими видами животных губки были отнесены к таксономической изоляции в пределах своего собственного типа. Однако, несмотря на их многочисленные общие характеристики, среди тысяч известных видов существует множество генетических различий. Четыре существующих класса внутри типа Porifera основаны на ключевых различиях в физиологии и среде обитания.
- Demospongiae : самый крупный и разнообразный из четырех классов, включающий более 70 процентов известных видов губок. У них мягкая, мясистая внешность, которая покрывает растянутую скелетную структуру, поддерживающую их вертикальный рост.
- Calcarea : Гораздо меньший класс из примерно 400 видов, характеризующихся спикулами на основе кальция, которые представляют собой твердые и заостренные наросты, служащие опорой и защитными структурами. Их спикулы имеют от 2 до 4 точек и состоят из карбонатов кальция, которые могут представлять собой либо арагонит, либо кальцит.
- Hexactinellida : Эти животные, также известные как «стеклянные губки», относятся к еще более редкому типу губок. У них часто есть 4 или 6 заостренных спикул, сделанных из соединений кремнезема, которые придают им уникальный внешний вид.
- Homoscleromorpha : самый маленький и самый примитивный из четырех классов. Эти губки могут растягиваться горизонтально и иметь простые биологические особенности по сравнению с видами других классов.
Губка: внешний вид животного
Учитывая тысячи различных видов внутри одного типа, неудивительно, что среди них огромное разнообразие размеров, форм и цветов. Большинство из них легко принять за кораллы или растения из-за их неподвижной природы и жесткой структуры. Они часто покрыты мягкой и мясистой внешностью, но их острые и твердые спикулярные скелеты могут быть уязвимы для хищников или из-за повреждений.
Для всех губок также характерно наличие отверстий вдоль их поверхности и каналов по всему телу. Поскольку у них нет внутренней системы кровообращения, эти поры позволяют воде проходить естественным путем, чтобы обеспечить кислород, ввести микроскопические частицы пищи и удалить отходы. Многие из этих животных имеют трубчатую форму с видимой в центре большой полостью, но они также могут вырастать в формы, похожие на деревья, веера или бесформенные капли. В зависимости от вида они также могут быть менее 1 дюйма в высоту или более 5 футов в высоту.
Губка на камне
Ник Хобгуд / Creative Commons
Губка: распространение, популяция и среда обитания животных
Представители типа Porifera встречаются во всех морях и океанах мира, а также в некоторых озерах и других пресноводных водоемах. . Подавляющее большинство из примерно 9000 известных видов живут исключительно в морской среде, причем от 100 до 200 обитают в пресноводных экосистемах. По оценкам некоторых исследователей, в отдаленных районах и глубоководных районах океана еще предстоит обнаружить многие тысячи видов губок.
Поскольку подавляющее большинство видов питаются планктоном и другими микроскопическими организмами, фильтруя окружающую воду, они предпочитают чистую и спокойную воду с минимальным загрязнением отложениями. Они часто прикрепляются к твердой поверхности, например к камням, рифам или даже к панцирям животных, но некоторые из них также могут отращивать достаточно длинные корни, чтобы прикрепляться к песку и другим рыхлым субстратам. Популяции обычно более разнообразны в тропическом климате по сравнению с умеренным и полярным.
губчатые хищники и добыча
Что поедает губки?
Отсутствие подвижности является серьезной биологической уязвимостью губок, что вынуждает развивать многие естественные механизмы защиты. Колючие спикулы на поверхности и выпущенные на окружающую местность помогают отпугивать морских звезд, морских ежей и других иглокожих, которые могут охотиться на губок. Потенциальные хищники могут включать различные виды насекомых, рыб, черепах и паразитов в зависимости от среды обитания. Губки также собирают и выращивают люди для различных коммерческих целей.
Чем питаются губки?
Большинство губок являются фильтраторами, что означает, что они питаются пассивно, потребляя микроскопические растения и животных из воды. Есть также некоторые виды, которые образуют симбиотические отношения с фотосинтетическими бактериями, что позволяет им получать энергию от солнечного света. Некоторые более мелкие губки пользуются своим размером и пассивной подвижностью, чтобы охотиться на других животных. Эти так называемые «скучные губки» прикрепляются к твердой поверхности моллюсков и разъедают раковину, чтобы охотиться на животное внутри. Моллюски, устрицы и другие моллюски являются основной целью, а также некоторые ракообразные.
Губка: размножение животных и продолжительность жизни
Половое размножение является типичным методом размножения, но некоторые виды могут также вести бесполое размножение. Большинство губок являются гермафродитами, что означает, что каждая особь имеет как мужские, так и женские клетки. При половом размножении губка выпускает яйца в воду, где они плавают, пока их не захватит другая губка, которая их оплодотворит. Губки могут вести обе деятельности одновременно или чередовать периоды выпуска и оплодотворения яиц. Средняя продолжительность жизни колеблется от менее 1 года до 20 лет, при этом некоторые виды способны выживать в течение многих столетий. Узнайте о самых долгоживущих животных здесь.
Оплодотворенные яйца высвобождаются в виде плавающих личинок, которые передвигаются с помощью слоя жгутиковых клеток. Как только они находят стабильную поверхность в подходящей среде, они прикрепляются и начинают метаморфозу в настоящую губку. Этот процесс включает в себя перемещение и трансформацию клеток по всему телу для облегчения развития специализированных функций.
Бесполое размножение часто является механизмом выживания, который позволяет губке выпускать небольшие колонии клеток. Этот процесс называется геммуляцией, и он позволяет вырождающемуся или умирающему взрослому высвобождать маленькие клоны, которые могли бы лучше жить в неблагоприятных условиях. Губки также обладают глубокими способностями к регенерации, поэтому маленькие кусочки могут превратиться в полностью выращенные клоны оригинала, если они оторваны от оригинала.
Использование губки в рыбной ловле и кулинарии
Аквакультура губки — это процветающая отрасль во многих регионах мира, преимущества которой заключаются в относительной простоте и небольших требованиях к материалам. Сельское хозяйство действительно зависит от благоприятных водных условий и последовательного управления для обеспечения продуктивных урожаев. Хотя они не используются людьми в качестве источника пищи, они находят практическое применение в купании, женской гигиене и в качестве источника биологических соединений. Биоактивные химические вещества обладают различными лечебными свойствами, включая противовоспалительный и противовирусный потенциал.
Просмотреть все 205 животных, названия которых начинаются с S
Об авторе
AZ Animals — это растущая команда экспертов по животным, исследователей, фермеров, защитников природы, писателей, редакторов и, конечно же, владельцев домашних животных, которые собраться вместе, чтобы помочь вам лучше понять животный мир и то, как мы взаимодействуем.
Часто задаваемые вопросы о губках (часто задаваемые вопросы)
Где можно найти губки?
Губки встречаются во всех видах водной среды. В зависимости от вида они могут процветать в прибрежных и приливных водах или в бездонных глубинах океанских желобов. Они встречаются в водах различной температуры, от мягкого тропического климата до холодных полярных вод. Подавляющее большинство живет в соленой воде, но есть некоторые виды, которые процветают в пресноводной среде.
Является ли губка животным?
Как члены королевства Animalia, губки технически являются животными. Они относительно уникальны среди животных из-за отсутствия у них органов, кровеносной или нервной системы. Биологи считают их сестринской группой всех других животных из-за того, как рано они разветвились в таксономическом дереве.
Двигаются ли губки?
У губок нет ног, и взрослые особи редко свободно плавают, но эмбрионы и маленькие губки могут передвигаться по воде. Некоторые виды также могут совершать очень медленные миграции, трансформируя и перемещая отдельные клетки вдоль своего основания.
Где живут губки?
Взрослые губки живут на субстратах или твердых поверхностях в водной среде. Большинство из них цепляются за скалы, рифы или другие твердые и устойчивые поверхности. Некоторые губки могут укореняться в рыхлом материале, таком как песок, в то время как другие цепляются за живые организмы, такие как черепахи, ракообразные или моллюски.
Что делает губка?
Большинство губок используют свои поры и внутренние каналы для фильтрации микроскопической пищи из окружающей воды. Они также выполняют важную структурную и симбиотическую роль в экосистемах, обеспечивая источник пищи и убежище для многих других видов животных.
Что едят губки?
Большинство губок питаются планктоном — микроскопическими растительными и животными организмами, плавающими в воде. Некоторые виды получают выгоду от фотосинтеза за счет симбиоза с бактериями, а некоторые виды губок фактически выживают, охотясь на моллюсков и ракообразных.
К какому королевству принадлежат Губки?
Губки относятся к царству Animalia.
К какому классу относятся губки?
Губки относятся к классу Demospongiae.
К какому типу относятся губки?
Губки относятся к типу Porifera.
Какое покрытие имеют губки?
Губки покрыты пористой кожей.
Какие хищники губки?
К хищникам губок относятся рыбы, черепахи и иглокожие.
Какой интересный факт о губках?
Более 9000 известных видов губок!
Какое научное название губки?
Научное название губки — Demospongiae.
Что является отличительной чертой Губки?
Губки имеют поверхностные поры.
Каков срок службы губки?
Губки могут жить от 15 до 30 лет.
Какая самая большая угроза для Губки?
Самая большая угроза губке — изменение климата и разрушение среды обитания.
Сколько губок осталось в мире?
Размер популяции Губки неизвестен.
Источники
- Дэвид Берни, Дорлинг Киндерсли (2011) Животное, Полное визуальное руководство по дикой природе мира
- Том Джексон, Лоренц Букс (2007) Всемирная энциклопедия животных
- Ричард Маккей, University of California Press (2009) Атлас исчезающих видов
- Дэвид Берни, Дорлинг Киндерсли (2008) Иллюстрированная энциклопедия животных
- Дорлинг Киндерсли (2006) Дорлинг Киндерсли Энциклопедия животных
- Рыболовство NOAA, доступно здесь: https://www.fisheries.noaa.gov/alaska/ecosystems/ trawling-effects-sponges-alaska
- Морское образовательное общество Австралии, доступно здесь: http://www. mesa.edu.au/porifera/
- Изучение нашей жидкой Земли, доступно здесь: https://manoa.hawaii. edu/exploringourfluidearth/биологические/беспозвоночные/филум-порифера
- Britannica, доступно здесь: https://www.britannica.com/animal/sponge-animal/Natural-history
- National Ocean Service, доступно здесь: https://oceanservice.noaa.gov/facts/sponge.html
- Википедия, доступно здесь: https://en.wikipedia.org/wiki/Sponge
Оценка потенциала губок (Porifera) как индикаторов концентраций растворенного Si в океане , и организмы-фильтраторы, принадлежащие к типу животных Porifera. Структура их тела проста, с различными видами клеток, специализирующихся на различных жизненных функциях и организованных в разные слои в соответствии с их функциями (Hooper and Van Soest, 2002; Van Soest et al., 2012; Maldonado, 2014). Их происхождение было отнесено к кембрию (Hooper and Van Soest, 2002; Xiao et al., 2005) или даже раньше, как предполагает открытие потенциальных биомаркеров губок (Love et al.
, 2009).; Инь и др., 2015; Голд и др., 2016).
В настоящее время выделяют четыре класса губок: Calcarea Carbonica, в которую входят виды с известковым скелетом; Hexactinellida, широко известные как стеклянные губки; Demospongiae, содержащий большинство современных губок; и Homoscleromorpha (Hooper and Van Soest, 2002; Morrow and Cádenas, 2015; Van Soest et al., 2017). К последним трем классам относятся виды с кремнистым скелетом, сцементированным органическими компонентами (т. е. коллагеноподобным белком, спонгином), а также виды, у которых отсутствует минеральный (т. армированные чужеродными неорганическими элементами (например, песком или остатками биогенного кремнезема из других организмов, включая губки; Hooper, Van Soest, 2002).
Благодаря сложным ферментативным и метаболическим процессам (Wang et al., 2011a,b, 2012a) кремнистые виды Hexactinellida, Demospongiae и Homoscleromorpha поглощают Si (преимущественно присутствующий в виде кремниевой кислоты, H 4 SiO 4 и условно обозначаемый в виде растворенного кремнезема; dSi) из окружающей их воды для выделения отдельных структур, известных как спикулы, которые обеспечивают опору скелета для органических компонентов губки (Uriz et al. , 2003). У некоторых видов гексактинеллид и демогубок скелетные структуры массивные и гиперкремнистые, каменистой консистенции, могут образовывать агломераты и рифоподобные образования (Conway et al., 2001; Uriz et al., 2003; Leys et al., 2004; Maldonado et al., 2015a,b) или «губки» (Klitgaard and Tendal, 2004). Губки этих групп могут играть важную роль в глобальном круговороте кремния из-за их широкого распространения и высокого потенциала сохранения (Maldonado et al., 2005, 2010).
Поглощение диоксида кремния и окварцевание губками регулируется наличием dSi в окружающей воде, а поглощение dSi соответствует ферментативной (Михалис-Ментен) кинетике с максимальной скоростью поглощения только выше 100–200 мкМ dSi (Reincke and Barthel, 1997; Мальдонадо и др., 1999, 2011). Эти значения выше, чем типичные концентрации в большей части современного океана, что позволяет предположить, что механизмы поглощения губками dSi развились в водах с более высоким dSi (Maldonado et al., 2015a), а dSi является ограничивающим фактором для живых губок (Maldonado et al. , 2011). Мальдонадо и др. (1999) также показал, что секреция определенных типов спикул регулируется порогами dSi, и предположил, что экземпляры этих спикул в ископаемых губках отражают среду, изобилующую dSi. Эффективные механизмы утилизации dSi развились у губок (Müller et al., 2003; Schröder et al., 2004; Wang et al., 2012a,b) и связаны с присутствием силикатеина, фермента, который способен контролировать и катализировать внутриклеточное отложение dSi из ненасыщенной среды, а также белки, активно транспортирующие dSi через клеточные мембраны (Schröder et al., 2004; Marron et al., 2016).
Дополнительная информация об изменениях формы или размера спикул губок или о нарушении спикулогенеза (изменения формы, атрофии или другие пороки развития спикул) в связи с концентрацией dSi разбросана по всей литературе (Jørgensen, 1944; Hartman, 1958; Stone, 1970; Yourassowsky and Rasmont, 1984; Zea, 1987; Bavestrello et al., 1993; Rützler and Smith, 1993; Schönberg and Barthel, 1997; Mercurio et al. , 2000; Valisano et al., 2012; Cádenas and Rapp. , 2013). В целом эти исследования показывают, что доступность dSi является фактором, который может влиять на морфологию, состав или размер спикул у некоторых видов.
Если dSi влияет на морфологию и/или морфометрию спикул и учитывая, что они являются основными признаками, используемыми для классификации кремнистых губок (Hooper and Van Soest, 2002), некоторые таксоны или группы таксонов могут быть характерны для местообитаний с особым условия dSi. Вероятно, они могут иметь разные механизмы адаптации и, следовательно, по-разному влиять на цикл кремния в океане. Важным первым шагом к использованию губок в качестве индикаторов является выявление и количественная оценка любых причинно-следственных связей между современным распространением губок и лежащими в их основе экологическими факторами; это важнейший первый шаг в разработке палеоэкологических прокси (например, Birks, 1995).
Пространственное распространение Porifera было изучено в местном и региональном масштабе и связано с различными факторами окружающей среды. Различия в видовом составе губок между мелководьем и глубоководьем признавались десятилетиями. Де Лаубенфельс (1936) признал, что пространственное распространение мелководных губок ограничено сильными барьерами окружающей среды (например, глубиной, светом или температурой). Совсем недавно пространственное распределение морских губок в местных масштабах (например, 10 км; коралловые рифы, проливы или заливы) коррелировали с геоморфологическими особенностями (Przeslawski et al., 2014), свойствами отложений, глубиной, расстоянием до побережье, доступность питательных веществ (включая dSi), проникновение света, гидродинамику (Huang et al., 2011), глубоководные течения (Cárdenas and Rapp, 2015) и биотические факторы, такие как хищничество или конкуренция за пространство (Huang et al., 2011). ; Pawlik et al., 2015; Slattery and Lesser, 2015). Как правило, эти наблюдения воспроизводятся в региональном или более широком масштабе (например, >100 км; моря или океаны). Здесь распределение губок было связано с глубиной, температурой и доступностью света (Vacelet, 1988; Финкс и Кит Ригби, 2003 г . ; Лейс и др., 2004; Downey et al., 2012) или в биогеографические регионы (Hooper and Lévi, 1994; Van Soest, 1994; Van Soest et al., 2012), ограниченные основными абиотическими факторами и характерными морскими биотами (например, Spalding et al., 2007). ). Интересно, что результаты прогностических моделей с использованием физико-химических переменных окружающей среды (Huang et al., 2011) и региональных исследований (Howell et al., 2016) показывают, что температура, глубина и состояние питательных веществ, включая dSi, являются основными факторами, контролирующими распространение губок.
Видовой состав губок и морфология спикул в осадочной летописи могут дать представление о dSi во время их образования. Реконструкции палео-dSi уделяется все больше внимания, отчасти из-за связи между долгосрочными биогеохимическими циклами углерода и кремния (Conley et al., 2000; Frings et al., 2016). На сегодняшний день реконструкция dSi с использованием губок достигается путем изучения морфометрических взаимоотношений пресноводных спикул с помощью градиентов dSi (Kratz et al. , 19).91). Совсем недавно при анализе изотопов кремния спикул губок использовалось наблюдение, согласно которому фракционирование стабильных изотопов кремния между окружающим dSi и биогенным Si в спикулах является функцией концентрации dSi в окружающей среде (Hendry et al., 2010, 2011; Wille и др., 2010; Хендри и Робинсон, 2012; Фонторбе и др., 2016, 2017). Разработка дополнительного палеоэкологического подхода, основанного на комплексах таксонов, могла бы значительно расширить понимание изменений dSi в океанах прошлого и настоящего. Оценивая распределение таксонов губок вдоль градиента dSi, можно идентифицировать определенные сообщества видов, чтобы получить представление о прошлой динамике Si в океане и прибрежной среде.
В этой работе мы исследуем распределение губок вдоль градиентов dSi, чтобы проверить, являются ли сообщества губок функцией dSi, и оценить достоверность таксономии губок как палеоэкологического инструмента для определения палеоконцентраций dSi. Для этого мы извлекли и объединили информацию из общедоступных океанографических баз данных и баз данных по биологическому разнообразию, чтобы оценить условия dSi, в которых губки встречаются в мировом океане. Поскольку dSi коррелирует с глубиной в современном океане, мы также исследуем распределение таксонов в зависимости от глубины. В целом мы обнаруживаем, что, хотя есть намеки на взаимосвязь между ассоциацией и dSi, губки недостаточно дифференцированы в зависимости от наличия dSi, чтобы четко указать конкретные концентрации dSi, что имеет значение для реконструкции цикла кремния в океане.
Материалы и методы
Источники данных
Мы использовали записи о губках, хранящиеся в Глобальном информационном центре по биоразнообразию (GBIF), и данные климатологии с координатной сеткой по химическому составу океана, интерполированные на основе измерений, полученных из Эксперимента по циркуляции Мирового океана (WOCE) (Gouretski and Колтерманн, 2004). Это доступно с разрешением сетки по горизонтали 0,5° и с 40 неравномерно расположенными вертикальными слоями. GBIF — это международная открытая инфраструктура данных и крупнейшая в мире база данных по биоразнообразию, которая обеспечивает свободный доступ к миллионам записей о биоразнообразии (http://www. gbif.org/what-is-gbif). GBIF объединяет данные из различных источников разного объема, качества и таксономической точности (дополнительную информацию см. на сайте www.gbif.org). WOCE — это глобальный ресурс данных Национального управления океанических и атмосферных исследований США, в конечном счете основанный на наблюдениях с кораблей, буев, дрифтеров и спутников. Мы опросили GBIF через онлайн-портал (http://www.gbif.org/occurrence) и загрузили 323,953 записи о Porifera с географической привязкой доступны в ноябре 2016 г. (загрузка GBIF Occurrence: https://doi.org/10.15468/dl.viufhq). Таксономическая классификация видов в этих записях соответствует классификации Всемирной базы данных Porifera (WPD, http://www.marinespecies.org/porifera/news.php?p=show&id=4893). В анализе использовались только записи с соответствующей глубиной (143 275; 44%). Палеонтологические записи и пресноводные таксоны, включенные в набор данных, также были удалены (830 и 192 записи соответственно). Чтобы проверить данные геолокации GBIF, мы сравнили глубину воды, связанную с каждой записью, с моделью глобального рельефа ETOPO1 1 угловая минута (Amante and Eakins, 2009). ). Поскольку оба набора данных содержат данные о глубине разного качества и разрешения, записи, которые расходились более чем на 500 м, были отброшены. Согласие было воспринято как признак уверенности в данных о глубине. Это 500-метровое пороговое значение было выбрано (довольно произвольно), чтобы учесть (i) возможность крутой батиметрии в пространственных масштабах ниже, чем у ETOPO1, и (ii) включение драгных проб, где дана только начальная или конечная точка траления. Этот процесс контроля качества привел к исключению 5190 (около 3,6%) точек данных, которые в некоторых случаях расходились на> 6000 м (см. Дополнительный рисунок 1). Кроме того, все оставшиеся записи, помеченные в GBIF как имеющие «нечисловую» или «неметрическую» глубину в исходном наборе данных (13 317 и 5 060 соответственно), были удалены, чтобы избежать проблем с преобразованием.
Информация на уровне класса и порядка, отсутствующая в некоторых записях GBIF, хотя был указан более низкий таксономический ранг семейства или рода, была добавлена после WPD. GBIF содержит смесь надежной и недостоверной таксономической информации (Van Soest et al., 2012) и включает записи с таксономической идентификацией, полученной из фотографий и/или видеозаписей, которые, например, Leys et al. (2004) определили как источник неопределенности. Мы предположили, что таксономические идентификации, включенные в записи GBIF на уровне отряда и семейства, более точны, чем таксономические идентификации более низких уровней (родов и видов), которые обычно требуют большей таксономической экспертизы. По этой причине мы ограничиваем наш анализ уровнем семейства и выше (порядок и класс). Наш анализ может быть улучшен и расширен до более высокого таксономического разрешения с повышением надежности таксономической идентификации записей GBIF.
Окончательный набор данных для этого исследования включал 62 360 дискретных глобально распределенных записей (рис. 1). Из них 2905 относятся только к классу, 890 — только к порядку (31 отряд), 2086 — к семейству (128 различных семейств), 24 525 — к роду (555 различных родов), а остальные (31 955 ) на виды (2758 различных видов) или подвиды. Для сравнения, Всемирная база данных Porifera (Van Soest et al., 2017) в настоящее время распознает 32 существующих отряда, 149существующих семейств и 740 существующих родов, что означает, что мы захватываем большую часть всего глобального биоразнообразия губок.
Рисунок 1 . Концентрации кремнезема, растворенного в океане, в придонных водах (т. е. на морском дне). Серые точки указывают местонахождение отдельных записей о губках, включенных в эту рукопись из GBIF.
Для каждого отдельного наблюдения губки в GBIF данные о температуре океана, солености и концентрации питательных веществ (нитратов, фосфатов и dSi) были извлечены из WOCE (Guretski and Koltermann, 2004) из коробки, в которой находится зарегистрированное местонахождение губки. Предоставляется сводка данных, включая подсчеты, максимумы, минимумы и средние значения для каждой переменной по таксонам для каждого из семейств, представленных в наборе данных (дополнительная таблица 1).
Влияние методов отбора проб и усилий по отбору проб
Концентрации dSi в придонных водах океана и местонахождение записей о губках из набора данных показаны на рисунке 1. которые мы пытаемся учесть в нашем анализе (см. ниже). Однако могут присутствовать и другие систематические ошибки выборки. Например, пробы в глубоководных местообитаниях обычно отбирают тралами или драгами. Обычно они ловят многочисленные, крупные или твердые экземпляры, но группы, представленные мелкими или хрупкими особями (например, Cladorhizidae и Calcarea Carbonica), часто не обнаруживаются (Raff et al., 19).94) и, следовательно, может быть недопредставлен. В настоящее время невозможно количественно оценить масштабы и значимость этого смещения.
Мы исследовали влияние очень интенсивного отбора проб в небольших регионах, сначала рассмотрев таксоны как присутствующие или отсутствующие только для каждой ячейки в наборе данных WOCE, и рассчитав глобальное распределение вдоль градиента окружающей среды с использованием 20 равноотстоящих интервалов для каждой переменной (показано для dSi на рисунке 2A). Агрегация присутствия/отсутствия лишь незначительно повлияла на распределение таксонов по градиентам окружающей среды (рис. 2B), поэтому мы работаем с полным (т. е. неагрегированным) набором данных. Чтобы грубо устранить смещение выборки в сторону континентальных шельфов и мелководных районов в записях GBIF, мы нормализовали распределения таксонов к общему распределению Porifera (рис. 2), чтобы получить набор распределений, которые описывают относительную важность отдельных таксонов для общей популяции Porifera вдоль любого заданного градиента окружающей среды. Этот подход неявно предполагает, что выборка и идентификация записей Porifera не смещены в сторону одной группы таксонов и не подходят, например, для калибровки экологических предпочтений отдельных таксонов губок. Однако цель здесь не в том, чтобы оценить истинное распределение таксонов губок в мировом океане, а в том, чтобы исследовать относительные различия среди таксонов. В дополнительной таблице 1 мы показываем сводку нескорректированных данных.
Рисунок 2. (A) Глобальное распределение записей GBIF, агрегированных как значения присутствия/отсутствия в полях 0,5° набора данных WOCE. (B) Глобальное распределение записей GBIF без поправки на систематическую ошибку выборки.
Скрипичные графики
Для многочисленных таксонов (более 100 записей в GBIF) были построены скрипичные графики нормированных данных в зависимости от основных экологических градиентов на разных уровнях таксономической классификации (класс, отряд и семейство) и составляют основу дальнейшего обсуждения. Там, где это применимо, мы включаем виды губок (далее именуемые «литистидами») с сильно окварцеванным скелетом, которые ранее относились к полифилетическому отряду Lithistida (Pisera and Lévi, 2002), но теперь разделены между неродственными отрядами класса Demospongiae (Cárdenas et al. др., 2012). На этих графиках ширина столбцов указывает относительный вклад таксонов в общее количество всех таксонов губок, зарегистрированных в GBIF при соответствующем значении переменной окружающей среды; обратите внимание, что они дополнительно стандартизированы для максимального вклада этих таксонов, поэтому не содержат никакой полезной информации об абсолютной численности любого данного таксона, что невозможно с данными GBIF «только для присутствия».
Результаты
Относительное частотное распределение таксонов губок в зависимости от dSi и глубины показано на рисунках 3–5. На уровне класса Hexactinellida встречаются в широком диапазоне значений dSi (0–180 мкМ; рис. 3A) и глубины (0–6000 м; рис. 3B). Губки с известковым скелетом демонстрируют аналогичное распределение, но вносят больший относительный вклад в общее количество таксонов губок при dSi < 100 мкМ и глубине < 3000 м. Остальные классы также находятся в том же диапазоне, однако их основной вклад в общее количество составляет менее 120 мкМ и менее 1000 м для кремнистых классов Homosclerophorida и Demospongiae.
Рисунок 3 . Скрипичные графики, показывающие распределение классов губок в градиенте dSi 90–140 (A) 90–141 и градиенте глубины 90–140 (B) 90–141.
В нашем анализе 19 из 32 полных отрядов губок представлены в GBIF как минимум в 100 блоках набора данных WOCE (рис. 4). Большинство из них встречаются преимущественно при dSi < 50 мкМ, в то время как другие группы демоспонгов (Tethyida, Suberitida Polymastiida и Haplosclerida), гексактинеллид (Hexactinosa и Lyssacinosida), гомосклерофорид и Clathrinida более равномерно распределены по градиенту dSi. Отряд гексактинеллид Amphidiscosida вносит наибольший вклад в сообщества губок в средах с dSi> 100 мкМ, а отряд известняковых Leucosolenida демонстрирует бимодальное распределение. На рис. 4Б показано распределение тех же порядков в зависимости от глубины. Сравнение двух графиков показывает, что отряды с относительно более частым появлением при более высоком dSi также представлены на глубинах более 2000 м. Отряды демогубок без кремнистого скелета (т.е. Dendroceratida, Verongiida – за исключением некоторых видов (Ehrlich et al., 2010) и Clionaida не обнаружены в базе данных GBIF на глубинах более 1000 м. Распределение литистид по градиенту глубины указывает на то, что эта группа вносит относительно постоянный вклад в общее количество губок при dSi ниже 50 мкМ и не обнаруживается на глубинах более 2500 м.
Рисунок 4 . Скрипичные графики, показывающие распределение порядков губок по градиенту dSi 90–140 (A) 90–141 и градиенту глубины 90–140 (B) 90–141. Показаны только заказы, представленные в GBIF не менее чем в 100 блоках набора данных WOCE. Цифры в скобках — это коды, присвоенные каждому из представленных заказов. Литистиды включают таксоны, отнесенные к этому таксону Писера и Леви (2002). Таксоны с серым фоном включают губки без кремнистых спикул. Цветовые коды, как на рис. 3.
На самом высоком уровне таксономического разрешения, которое мы рассматриваем, 43 семейства в пределах Porifera встречаются в 100 или более ячейках WOCE (для справки: в настоящее время в пределах Porifera признано 149 семейств; Van Soest et al., 2017). Их распределения следуют тем же тенденциям, что и показанные на уровне порядка, при этом некоторые семейства лучше представлены при dSi < 50 мкМ, некоторые примерно одинаково представлены вдоль градиента, некоторые встречаются как при высоком, так и при низком dSi, а другие с большей вероятностью будут обнаружены при dSi. > 100 мкМ (рис. 5А). Распределение этих семейств по градиенту глубины (рис. 5B) примерно соответствует градиенту dSi. Семейства, встречающиеся при высоком dSi, такие как Cladorhizidae, Polymastiidae и все гексактинеллиды, также, вероятно, обитают в глубоких водах. Некремнистые губки отрядов Verongiida, Dictyoceratida и Dendroceratida, за исключением Spongiidae (см. обсуждение ниже), встречаются в группе таксонов, встречающихся при dSi < 20 мкМ, вместе с семейством Clionidae (обычно губки, ассоциированные с кальцием). карбонатные субстраты), но и с семействами, имеющими кремнистый скелет (например, Petrosiidae, Dictyonellidae и Callyspongiidae). Они встречаются только на небольшой глубине (рис. 5А, В).
Рисунок 5 . Скрипичные графики, показывающие распределение семейств губок в градиенте dSi 90–140 (A) 90–141 и градиенте глубины 90–140 (B) 90–141. Показаны только семьи, представленные в GBIF не менее чем в 100 блоках набора данных WOCE. Цифры в скобках — это коды, присвоенные каждой семье. Таксоны с серым фоном включают губки без кремнистых спикул. Ключ к заказам, представленным на рисунке 4, включен. Цветовые коды как на рис. 3.
Обсуждение
Наш анализ обеспечивает количественную оценку диапазонов dSi таксонов обыкновенных губок и расширяет предыдущие знания, касающиеся их батиметрии (Vacelet, 1988; Maldonado and Young, 1998; Rapp et al. , 2011; Downey and Janussen, 2015; Hestetun et al. ., 2015). Распределение таксонов губок по градиенту глубины демонстрирует явное сходство с dSi, а это означает, что задача состоит в том, чтобы провести различие между ними.
Ниже мы обсудим роль доступности dSi и влияние других переменных, которые могут косвенно влиять на представленные здесь закономерности. Мы также обсудим, как эти данные могут способствовать прояснению роли губок в круговороте кремния в океане в геологическом прошлом, а также полезность данных о комплексах губок в качестве морского палеоэкологического инструмента.
Влияние наличия dSi и глубины среды обитания на распространение губок
Глубина является важным фактором, контролирующим распространение губок (Vacelet, 1988; Huang et al., 2011; Howell et al., 2016), хотя он действует косвенно через корреляцию с экологически значимые параметры (включая концентрацию dSi, а также свет, доступность пищи, температуру и т. д.). Очевидно, что либо dSi, либо глубина, либо и то, и другое играют важную роль, контролируя пространственное распределение таксонов губок (рис. 3–5). Наши результаты подтверждают, что Porifera неравномерно распределены ни по градиенту dSi, ни по глубине. Поскольку дифференциация вдоль двух градиентов аналогична, неясно, влияет ли наличие dSi, глубина среды обитания или другой неустановленный фактор (или комбинация факторов) на пространственное распределение губок. По крайней мере, в региональном масштабе глубина воды действует как суррогатная переменная для проникновения света, температуры и концентрации питательных веществ; все они взаимосвязаны и могут влиять на распределение губок (Vacelet, 1988). Другие факторы, связанные с глубиной, такие как пищевые ресурсы (т. е. твердые и растворенные вещества), турбулентность, пространственная конкуренция и хищничество, были предложены в качестве движущих сил структуры и распределения сообщества губок (Pawlik et al., 2015; Slattery and Lesser, 2015). .
Температура придонной воды и концентрация некоторых питательных веществ также были определены как важные переменные, контролирующие распространение губок в местных масштабах (Huang et al. , 2011; Howell et al., 2016). Однако распределение губок в зависимости от температуры, концентраций растворенного кислорода и питательных веществ (нитратов и фосфатов), извлеченных из WOCE (см. Дополнительный материал), показало, что распределение таксонов губок вдоль этих градиентов не сильно дифференцировано, в отличие от dSi. и градиенты глубины. Хотя мы не можем исключить эти факторы, тот факт, что глубина местообитания демонстрирует сходную картину распределения губок с dSi, позволяет предположить, что доступность dSi является важной движущей силой пространственного распределения некоторых групп губок, и подразумевает, что распределение губок как функция глубины просто отражает общая взаимосвязь между глубиной и [Si] (см. ниже; рис. 6). К сожалению, мы не можем строго определить направление причинно-следственной связи и исключить возможность того, что глубина просто действует как суррогат другой, пока не идентифицированной переменной.
Рисунок 6 . Кросс-плот средней глубины среды обитания и средних концентраций растворенного кремния в окружающей среде для всех семейств губок в базе данных GBIF. См. предыдущие рисунки для ключей к цветам. Все таксоны включены независимо от их численности, в отличие от рисунков 3–5, где применяется критерий встречаемости 100 ячеек WOCE. Данные агрегируются на уровне присутствия/отсутствия для отдельных ячеек WOCE. Серые точки на заднем плане — концентрации dSi в придонной воде океана (т. е. те же данные, что и на рис. 1).
В целом dSi океана коррелирует с глубиной (Sarmiento and Gruber, 2006), поскольку биогенный кремнезем, продуцируемый кремнистым планктоном на поверхности, постепенно растворяется по мере его погружения в толщу воды (хотя эта закономерность несколько осложняется морской опрокидывающей циркуляцией, которая смешивает водные массы разного возраста и, следовательно, накапливает dSi), а также из-за высокой продукции диатомовых водорослей из-за апвеллинга. Для дальнейшего изучения этого вопроса мы нарисовали dSi и глубину для всех семейств, включенных в наш набор данных на рисунке 6. Это подтверждает наличие положительной зависимости между dSi и глубиной, форма которой аналогична той, что для всего океана (рисунок 6).
Сообщества губок вдоль dSi и градиентов глубины
Наши данные показывают, что либо глубина, либо dSi играют основную роль, контролируя пространственное распределение некоторых таксонов (рис. 3–5). Ниже мы обсудим, какие из этих таксонов могут включать виды с различными требованиями к поглощению dSi и, следовательно, могут указывать на условия dSi.
Hexactinellida
Виды отряда Hexactinosa (например, Farrea occa, Aphrocallistesvastus и Heterochone calyx ), как известно, строят гиперкремниевые скелеты и формируют губчатые рифы, где dSi достаточно высок (около 40 мкМ), например, на западном побережье Канады (Leys et al., 2004; Conway et al., 2005; Whitney et al. и др., 2005; Чу и др., 2011). Считается, что появление этих рифообразующих видов контролируется совместным действием определенных океанографических, геоморфологических условий и биотических факторов, включая доступность dSi (Conway et al., 2005; Maldonado et al., 2015a). Hexactinosa (рис. 4) являются важными компонентами фауны губок на высоких dSi и глубине вместе с видами, принадлежащими Amphidiscosida, Aulocalycoida и Lyssacinosida (Rossellidae), у которых отсутствует диктионный (сросшиеся спикулы в трехмерном каркасе) скелет, пригодный для формирования рифов, но тем не менее имеют относительно больше dSi на единицу биомассы, чем губки из других классов губок (Bartel, 1995).
Demospongiae
Семейства Cladorhizidae и Polymastiidae относительно чаще встречаются в водах, богатых dSi (рис. 5А). Эти семейства также имеют широкий диапазон распространения вдоль градиентов глубины и, вероятно, могут быть обнаружены в глубоких водах (> 2000 м), где dSi также высок. Еще предстоит ответить, зависит ли распределение этих таксонов от dSi или от глубины. Подробные исследования механизмов поглощения dSi и скорости этих видов губок, живущих в различных условиях dSi, помогут ответить на этот вопрос и выявить виды, которые можно использовать в качестве индикаторов конкретных диапазонов dSi.
Оставшиеся кремнистые и некремнистые демогубки, скорее всего, обитают в местах обитания с низким dSi, хотя некоторые из них могут встречаться на больших глубинах с высоким dSi. Некоторые из семейств Dictyoceratida (например, Spongiidae), у которых отсутствует кремнистый скелет, скорее всего, встречаются на больших dSi и глубине (рис. 5). Это интересное наблюдение, поскольку принято считать, что представители этой группы обитают в тропических широтах и на мелководье (Vacelet, 1988), а у некоторых видов на расселение их личинок влияют низкие температуры и проникновение света (Maldonado, 2009).). Насколько нам известно, виды Spongiidae из батиальных глубин в таксономической литературе не описаны. Возможно, эти записи могут быть ошибочно идентифицированы, учитывая, что таксономия этих губок чрезвычайно сложна. Однако до независимой проверки этих записей Dictyoceratida мы предполагаем, что их документация в GBIF верна, и, следовательно, другие факторы, коррелирующие с глубиной (включая температуру, проникновение света, концентрацию нитратов или фосфатов), влияют на появление определенных видов этой группы.
Lithistids
Губки, включенные в эту полифилетическую (различное филогенетическое родство) группу, включают виды с сочлененными спикулами, называемыми десмами, которые у большинства видов образуют жесткий скелет (Pisera and Lévi, 2002). Большинство (93%) записей литистид относятся к отряду Tetractinellida, а наиболее распространенными семействами были Theonellidae (21%), Corallistidae (18%) и Scleritodermidae (17%). Согласно рисунку 4, таксоны литистид с большей вероятностью будут обнаружены при < 60 мкМ dSi, с наибольшей относительной частотой при < 20 мкМ и < 1000 м. Известно, что некоторые из них строят рифоподобные образования в местах обитания, где доступность dSi низка по сравнению с таковой, о которой сообщалось для рифоподобных строителей Hexactinosida (Maldonado et al., 2015a,b). Гиперкремнистые виды lithistid (например, Leiodermatium spp) также сосуществует с представителями губок без кремнистого скелета (т. е. Dictyoceratida, Dendroceratida и Verongiida) (Przeslawski et al. , 2014, 2015) в относительно неглубоких местообитаниях северной Австралии с концентрациями dSi <5 мкМ, подверженных воздействию сильные приливно-отливные течения, высокая мутность и значительная подвижность наносов (Przeslawski et al., 2011). Как было предложено Мальдонадо и соавт. (2015b), некоторые виды литистид, по-видимому, извлекают выгоду из отложения тяжелых отложений и моделей местной циркуляции, которые могут доставлять пищу и Si в импульсах. Остается ответить, как эти гиперокремненные скелеты литистид могут развиваться при более низком dSi и могут ли эти виды иметь более эффективные системы поглощения Si, чем другие таксоны, такие как Hexactinosida, которые, как правило, обнаруживаются при сравнительно более высоких dSi (рис. 4, 5). ).
Calcarea
Данные GBIF показывают, что губки класса Calcarea встречаются на глубине до 5000 м с пиками ниже 3000 м. Это подтверждает предыдущие выводы (Duplessis and Reiswig, 2000; Rapp et al., 2011) и позволяет предположить, что разнообразие группы на абиссальных глубинах выше, чем предполагалось ранее, и что иногда они способны расти в средах с более низким уровнем насыщения карбонатными минералами.
На пути к количественной оценке
Существует большое количество литературы, посвященной методам получения количественных реконструкций палеосреды на основе (микро) ископаемых комплексов в наземных, пресноводных и морских средах. Как резюмировано, например, в Telford and Birks (2005) или Juggins (2013), все больше осознается ограниченность этих подходов, даже в благоприятных ситуациях. Мы отказались от более количественного подхода по нескольким причинам, в том числе (i) данные GBIF собраны из разрозненных, несвязанных источников с разными стратегиями выборки, (ii) верхняя граница градиента dSi плохо представлена в данных, (iii) данные «только о присутствии», а не об истинной численности видов, что означает, что оптимальные таксоны не могут быть легко определены, (iv) у нас нет независимого набора данных для проверки модели, (v) разделение дисперсии предположительно будет указывать на высокую степень общей дисперсии между глубиной и dSi, и (vi) данные калибровки окружающей среды имеют трудно поддающийся количественной оценке шумовой компонент, полученный отдельно от данных GBIF с более низким пространственным разрешением.
В будущем истинная совокупность губок (с численностью таксонов) и набор данных переменных окружающей среды, собранный в регионе или регионах, где dSi не связан с глубиной, смогут преодолеть некоторые из этих трудностей. Соотношения изотопов кремния в губке все шире используются в качестве косвенного показателя концентраций dSi в прошлом (например, Fontorbe et al., 2017). Хотя и не получившие широкого распространения, подходы «взаимного диапазона» (Sinka and Atkinson, 1999), современные аналоговые методы (MAT; Lytle and Wahl, 2005) или искусственные нейронные сети (Racca et al., 2007) могут оказаться плодотворными палеоэкологическими дополнениями. Наконец, лабораторные и полевые эксперименты, тщательно разработанные для изучения влияния доступности dSi на физиологию и экологию губок, или подходы, направленные на использование морфометрии отдельных спикул (см. Kratz et al., 19).91) также может быть частью разрабатываемого «набора инструментов» палео-dSi.
Контролировала ли dSi распространение губки в прошлом?
Эволюция цикла кремния в океане привлекала внимание на протяжении многих десятилетий (например, Maliva et al. , 1989; Kidder and Tomescu, 2016). Преобладающий нарратив (Малива и др., 1989; Сивер, 1991; Киддер и Томеску, 2016) приводит доводы в пользу двух особенно важных событий. Первый – это переход от докембрийского океана, насыщенного по dSi, к малонасыщенному мезозойскому океану. Вторым было дальнейшее снижение где-то в меловом-палеогеновом периоде содержания dSi в средней океанской воде до уровня современного океана, обедненного dSi. Оба перехода приписываются биологическим инновациям — первый — эволюции окварцевания радиоляриями и губками, а второй — распространению диатомовых водорослей — и, следовательно, прогрессивной способности биологии удалять dSi из морской воды. В связи с этим долгосрочным снижением dSi в океане были предложены переходные отклонения запаса dSi в океане в ответ на периоды экстенсивного вулканизма (в частности, формирование крупных изверженных провинций, LIP) (Ritterbush et al., 2014, 2015; Kidder and Tomescu, 2016), или орогенез (например, Cermeño et al., 2015). Одним из доказательств, подтверждающих эти интерпретации, является изменение осадочных фаций, связанных с губками, в горных породах или отложениях (например, Kidder and Tomescu, 2016). Связь между численностью/распределением губок и dSi проистекает из неявного предположения, что кремнистые губки не могут развиваться при низком dSi (например, Ritterbush et al., 2015; Kidder and Tomescu, 2016). Киддер и Томеску (2016), например, интерпретируют ордовикское отступление губок от лагунных условий осадконакопления к более глубоким шельфам как отражение вызванного радиоляриями истощения dSi на поверхности океана. После середины ордовика окаменелости губок из периторальных и лагунных фаций, по-видимому, редки в летописи окаменелостей (см. Gammon et al., 2000) за возможное эоценовое исключение. Малива и др. (1989) утверждают, что губки в кайнозое теряют количественное значение в круговороте кремния в океане. В какой степени эти предполагаемые изменения численности и распределения губок обусловлены доступностью dSi?
Наш набор данных показывает, что все группы губок, из которых было отобрано достаточно образцов (т. Е. > 100 ящиков WOCE, см. Дополнительную таблицу 1), способны расти в средах с низким dSi. Действительно, по крайней мере в некоторых отдельных случаях таксоны кремнистых губок способны образовывать большие рифоподобные скопления при <10 мкМ dSi (Maldonado et al., 2015a,b). Это означает, что, хотя любое снижение доступности dSi могло быть фактором, способствующим исключению губок из среды мелководья, следует принимать во внимание и другие факторы, возможно, связанные с эволюцией и экологией конкурентов губок. Следовательно, присутствие любого данного таксона само по себе не может свидетельствовать о преобладающих концентрациях dSi.
Например, рифообразующие литигистиды, использовавшиеся в качестве индикаторов большей доступности dSi в прошлых средах (например, Corsetti et al., 2015; Ritterbush et al., 2015), в нашем наборе данных встречаются чаще <20 мкМ и < 2500 м. В настоящее время нет никаких доказательств того, что они не могли жить в подобных условиях в геологическом прошлом (Pisera, 1997). Расширение некоторых групп губок после триасово-юрского вымирания, скорее, является следствием большей доступности пригодных экологических ниш, освободившихся после распада известковых беспозвоночных в этот период (Ritterbush et al. , 2014, 2015).
Наконец, важно отметить, что, как правило, большинство кремнистых и некремнистых губок плохо представлены в летописи окаменелостей, поскольку их скелеты легко разрушаются. Виды с жестким скелетом, например литистиды, некоторые гексактинеллиды и ископаемые губки с базальным известковым скелетом (т.
Выводы
Здесь мы показываем, что распределение губок, классифицированных на уровне семейства, связано с концентрацией dSi в окружающей среде или глубиной среды обитания, или и тем, и другим. Характер распределения по концентрациям dSi или глубине местообитания указывает на то, что большинство таксонов, включая таксоны класса Calcarea без кремнистого скелета, встречаются в широком диапазоне dSi (0–180 мкМ) и глубине (0–~5500 м). Это также показывает, что большинство демогубок без автохтонного кремнистого скелета встречается при dSi <20 мкМ и небольших глубинах, но вместе с группами, у которых скелет состоит из кремнистых спикул. Демогубки с жестким кремнистым скелетом, традиционно известные как литистиды, по-видимому, не связаны с глубокими средами, богатыми dSi, что указывает на то, что этот вид губок мог выработать особенно эффективные механизмы для включения и накопления dSi в своих скелетах. С другой стороны, гексактинеллиды, также создающие прочный и жесткий кремнистый скелет, распределяются вдоль градиента dSi, но с большей вероятностью присутствуют в глубоких и богатых кремнеземом средах. Гексактинеллиды разделяют такую среду с некоторыми демоспонгами, равномерно распределенными по dSi и градиенту глубины, и в основном из отрядов Cladorhizidae и Polymastiida.
Группы с перекосом в сторону низкого или высокого dSi могут включать виды, характерные для конкретных условий содержания кремния в окружающей среде. Поскольку уникального или четкого набора таксонов выявить не удалось, требуется дополнительная информация о скорости поглощения Si для возможных показательных видов, чтобы окончательно связать их пространственное распределение с уровнями dSi. Можно было бы ожидать большей дифференциации вдоль градиентов окружающей среды, если рассматривать губки на уровне родов или даже видов, хотя мы отмечаем, что некоторые роды (например, Geodia ) очень космополитичны. Точно так же изучение механизмов поглощения dSi видами губок, встречающимися в широком диапазоне dSi и глубин, может предоставить важную информацию об адаптационном потенциале таксонов губок.
Связи между обилием губок в осадочных записях и современными концентрациями dSi следует интерпретировать с осторожностью, учитывая, что эти организмы явно приспособлены к жизни в диапазоне концентраций dSi и глубинных местообитаний, как видно из наших результатов. Другие биотические (например, конкурентные взаимодействия) и абиотические факторы (например, проникновение света, температура, доступность питательных веществ и пищи), некоторые из которых также связаны с глубиной, могут быть важными факторами в распространении современных губок и, вероятно, не менее важны. в палеосредах.
Вклад авторов
Все перечисленные авторы внесли существенный, непосредственный и интеллектуальный вклад в работу и одобрили ее для публикации. BA и PF являются совместными старшими авторами.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Это исследование финансировалось за счет гранта Фонда Кнута и Элис Валленберг округу Колумбия. Дмитрий Шигель, GBIF, и Меррик Экинс, Музей Квинсленда, предоставили ценные комментарии и предложения, связанные с данными, загруженными с портала GBIF. Мы благодарим всех за предложения и исправления рецензентов.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fmars.2017.00373/full#supplementary-material
Ссылки
Amante, C. и Икинс, Б.В. (2009). Etopo1 1 Модель глобального рельефа угловых минут: процедуры, источники данных и анализ . Технический меморандум NOAA NESDIS NGDC-24. Боулдер, Колорадо: Национальный центр геофизических данных, NOAA.
Бартель, Д. (1995). Состав тканей губок из моря Уэдделла, Антарктида: на костях мало мяса. Мар. Экол. прог. сер. 123, 149–153. doi: 10.3354/meps123149
Полный текст CrossRef | Google Scholar
Бавестрелло Г. , Бонито М. и Сара М. (1993). Изменение содержания кремнезема и размера игл в течение годового цикла у Chondrilla nucula Schmidt (Porifera, Demospongiae) в Лигурийском море. Науч. 90 525 марта 57, 421–425.
Google Scholar
Биркс, HJB (1995). «Количественные реконструкции палеоокружающей среды», в Статистическое моделирование четвертичных научных данных , редакторы Д. Мэдди и Дж. С. Брю (Кембридж, Ассоциация четвертичных исследований), 161–254.
Карденас, П., Перес, Т., и Бури-Эно, Н. (2012). «Глава вторая — систематика губок сталкивается с новыми проблемами», в Advances in Marine Biology , редакторы М. А. Бесерро, М. Дж. Урис, М. Мальдонадо и X. Турон (Лондон: Academic Press), 79–209..
Карденас, П., и Рапп, Х. Т. (2013). Нарушенный спикулогенез у глубоководных Geodiidae (Porifera, Demospongiae), растущих на мелководье. Инвертебр. биол. 132, 173–194. doi: 10.1111/ivb.12027
Полный текст CrossRef | Google Scholar
Карденас, П. , и Рапп, Х. Т. (2015). Демогубки из Северного Срединно-Атлантического хребта проливают больше света на разнообразие и биогеографию глубоководных губок Северной Атлантики. J. Mar. Biol. доц. Великобритания 95, 1475–1516. дои: 10.1017/S0025315415000983
Полнотекстовая перекрестная ссылка | Google Scholar
Черменьо П., Фальковски П. Г., Ромеро О. Э., Шаллер М. Ф. и Валлина С. М. (2015). Континентальная эрозия и кайнозойский подъем морских диатомей. Проц. Натл. акад. науч. США . 112, 4239–4244. doi: 10.1073/pnas.1412883112
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чу, Дж., Мальдонадо, М., Яхель, Г., и Лейс, С. (2011). Стеклянная губка действует как силиконовая раковина. Мар. Экол. прог. сер. 441, 1–14. doi: 10.3354/meps09381
Полный текст CrossRef | Google Scholar
Конли Д. Дж., Столнаке П., Питканен Х. и Виландер А. (2000). Перенос и удержание растворенного силиката реками в Швеции и Финляндии. Лимнол. океаногр. 45, 1850–1853 гг. doi: 10.4319/lo.2000.45.8.1850
CrossRef Full Text | Google Scholar
Конвей К.В., Крауттер М., Барри Дж.В. и Нойвайлер М. (2001). Гексактинеллидные губчатые рифы на континентальном шельфе Канады: уникальное «живое ископаемое». Геофиз. Можно. 28, 71–78. Доступно в Интернете по адресу: https://journals.lib.unb.ca/index.php/GC/article/view/4076
Google Scholar
Conway, K.W., Krautter, M., Barrie, J.V., Whitney, F. , Thomson, R.E., Reiswig, H., et al. (2005). Губчатые рифы в бассейне Королевы Шарлотты, Канада: меры контроля за распространением, ростом и развитием (Берлин: Springer-Verlag Berlin), 605–621.
Google Scholar
Корсетти Ф. А., Риттербуш К. А., Ботьер Д. Дж., Грин С., Ибарра Ю., Ягер Дж. А. и др. (2015). Изучение палеоэкологических последствий распада суперконтинента: губки очищаются в ранней юре. Осадок. Рек. 13, 4–10. Доступно в Интернете по адресу: https://www. sepm.org/CM_Files/SedimentaryRecord/SedRecord13-2%235.pdf
Google Scholar
De Laubenfels, M.W. (1936). Сравнение мелководных губок у тихоокеанского конца Панамского канала с губками на карибском конце. Проц. Натл США. Мус. 83, 441–464. doi: 10.5479/si.00963801.83-2993.441
CrossRef Full Text
Downey, R.V., Griffiths, H.J., Linse, K., and Janussen, D. (2012). Разнообразие и закономерности распространения губок высоких южных широт. PLoS ONE 7:e41672. doi: 10.1371/journal.pone.0041672
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Дауни, Р. В., и Януссен, Д. (2015). Новые взгляды на фауну абиссальных губок Курило-Камчатской равнины и желоба (северо-западная Пацифика). Deep Sea Res. II Топ. Стад. океаногр. 111, 34–43. doi: 10.1016/j.dsr2.2014.08.010
CrossRef Полный текст | Google Scholar
Дюплесси К. и Рейсвиг Х. (2000). Описание новой глубоководной известковой губки ( Porifera: Calcarea ) из Северной Калифорнии. Упак. науч. 54, 10–14. Доступно на сайте: http://hdl.handle.net/10125/1593
Google Scholar
Эрлих Х., Саймон П., Каррильо-Кабрера В., Баженов В.В., Боттинг Дж.П., Илан, М. и др. (2010). Взгляд на химию биологических материалов: недавно обнаруженные биокомпозиты кремний-арагонит-хитин в демогубках. Хим. Мат. 22, 1462–1471. doi: 10.1021/cm
Полный текст CrossRef | Академия Google
Финкс, Р. М., и Кит Ригби, Дж. (2003). «Географическое и стратиграфическое распространение», в Трактате по палеонтологии беспозвоночных. Часть E (пересмотренная), Porifera , Vol. 2, изд. Р.Л. Кеслер (Боулдер, Колорадо; Лоуренс, Канзас: Геологическое общество Америки и Канзасский университет), 275–296.
Фонторбе Г., Фрингс П., Де Ла Роша К.Л., Хендри К.Р. и Конли Д.Дж. (2016). Кремний истощал Северную Атлантику со времен палеогена: свидетельство изотопов кремния губок и радиолярий. Планета Земля. науч. лат. 453, 67–77. doi: 10.1016/j. epsl.2016.08.006
CrossRef Full Text | Google Scholar
Fontorbe, G., Frings, P.J., De La Rocha, C.L., Hendry, K.R., Carstensen, J., and Conley, D.J. (2017). Обогащение растворенного кремнезема в глубокой экваториальной части Тихого океана в эоцен-олигоцене. Палеоокеанография 32, 848–863. doi: 10.1002/2017PA003090
Полный текст CrossRef | Google Scholar
Фрингс П. Дж., Клайманс В., Фонторбе Г., Де Ла Роча К. Л. и Конли Д. Дж. (2016). Континентальный цикл кремния и его влияние на баланс изотопов кремния в океане. Хим. геол. 425, 12–36. doi: 10.1016/j.chemgeo.2016.01.020
CrossRef Полный текст | Google Scholar
Gammon, P.R., James, N.P., and Pisera, A. (2000). Эоценовые спикулиты и спонголиты на юго-западе Австралии: не глубокие, не полярные, а неглубокие и теплые. Геология 28, 855–858. doi: 10.1130/0091-7613(2000)28<855:ESASIS>2.0.CO;2
CrossRef Full Text | Google Scholar
Голд, Д. А., Грабенстаттер, Дж. , де Мендоса, А., Рисго, А., Руис-Трилло, И., и Саммонс, Р. Э. (2016). Стероловый и геномный анализы подтверждают гипотезу биомаркера губки. Проц. Натл. акад. наука . США . 113, 2684–2689. doi: 10.1073/pnas.1512614113
Полный текст CrossRef | Google Scholar
Гурецкий В.В. и Колтерманн К.П. (2004). Глобальная гидрографическая климатология WOCE. 35/2004. Berichte des Bundesamtes fur Seeschifffahrt und Hydrographie.
Google Scholar
Hartman, WD (1958). Естественная история морских губок на юге Новой Англии. Музей Пибоди Нац. История 12, 1–155.
Google Scholar
Хендри, К. Р., Георг, Р. Б., Рикаби, Р. Э. М., Робинсон, Л. Ф., и Холлидей, А. Н. (2010). Глубинные питательные вещества океана во время последнего ледникового максимума выведены из изотопного состава кремния губок. Планета Земля. науч. лат. 292, 290–300. doi: 10.1016/j.epsl.2010.02.005
CrossRef Full Text | Google Scholar
Хендри, К. Р., Ленг, М. Дж., Робинсон, Л. Ф., Слоан, Х. Дж., Блаштьян, Дж., Рикаби, Р. Э. М., и др. (2011). Изотопы кремния в антарктических губках: межлабораторное сравнение. Антаркт. науч. 23, 34–42. doi: 10.1017/S0954102010000593
Полный текст CrossRef | Google Scholar
Хендри, К. Р., и Робинсон, Л. Ф. (2012). Взаимосвязь изотопного фракционирования кремния в губках и концентрации кремниевой кислоты: современные и керновые исследования биогенного опала. Геохим. Космохим. Акта 81, 1–12. doi: 10.1016/j.gca.2011.12.010
Полный текст CrossRef | Google Scholar
Hestetun, JT, Fourt, M., Vacelet, J., Boury-Esnault, N., and Rapp, H.T. (2015). Cladorhizidae ( Porifera, Demospongiae, Poecilosclerida ) глубоководных районов Атлантического океана, собранных во время круизов Ифремера, с биогеографическим обзором атлантических видов. J. Mar. Biol. доц. Великобритания 95, 1311–1342. doi: 10.1017/S0025315413001100
Полный текст CrossRef | Google Scholar
Хупер, Дж. Н. А., и Леви, К. (1994). «Биогеография индо-западно-тихоокеанских губок: microcionidae, raspailiidae, axinellidaem», в Sponges in Time and Space , eds RWM Van Soest, TMG Van Kempen и JC Braekman (Rotterdam: Balkema), 191–212.
Хупер, Дж. Н. А., и Ван Соест, Р. В. М. (ред.). (2002). Systema Porifera: Руководство по классификации губок. Нью-Йорк, штат Нью-Йорк: Kluwer Academic/Plenum Publishers.
Google Scholar
Хауэлл К.-Л., Пьешо Н., Дауни А.-Л. и Кенни А. (2016). Распределение скоплений глубоководных губок в Северной Атлантике и последствия для их эффективного пространственного управления. Deep Sea Res. Я океаногр. Рез. Пап. 115 (Прил. С), 309–320. doi: 10.1016/j.dsr.2016.07.005
Полный текст CrossRef | Google Scholar
Хуанг З., Брук Б. и Ли Дж. (2011). Эффективность прогностических моделей в морской бентической среде на основе прогнозов распределения губок на континентальном шельфе Австралии. Экол. Поставить в известность. 6, 205–216. doi: 10.1016/j.ecoinf.2011.01.001
CrossRef Полный текст | Google Scholar
Jørgensen, CB (1944). О спикулообразовании Spongilla lacustris (L.): зависимость спикулообразования от содержания растворенной и твердой кремниевой кислоты в среде . Копенгаген: Э. Мунксгаард (Бьянко Лунос Богтриккери).
Джаггинс, С. (2013). Количественные реконструкции в палеолимнологии: новая парадигма или больная наука? Кв. науч. Ред. 64, 20–32. doi: 10.1016/j.quascirev.2012.12.014
CrossRef Полный текст | Google Scholar
Киддер Д.Л. и Томеску И. (2016). Биогенный кремень и ордовикский цикл кремнезема. Палеогеогр. Палеоклимат. Палеоэколь. 458, 29–38. doi: 10.1016/j.palaeo.2015.10.013
Полнотекстовая перекрестная ссылка | Google Scholar
Клитгаард А.Б. и Тендал О.С. (2004). Распространение и видовой состав массовых скоплений крупных губок в северо-восточной Атлантике. Прог. океаногр. 61, 57–98. doi: 10. 1016/j.pocean.2004.06.002
CrossRef Полный текст | Google Scholar
Крац Т.К., Фрост Т.М., Элиас Дж.Э. и Кук Р.Б. (1991). Реконструкция регионального 12000-летнего упадка кремнезема в озерах с помощью ископаемых спикул губок. Лимнол. океаногр. 36, 1244–1249. doi: 10.4319/lo.1991.36.6.1244
CrossRef Full Text | Google Scholar
Лейс С.П., Уилсон К., Холетон К., Рейсвиг Х.М., Остин У.К. и Танниклифф В. (2004). Закономерности распространения стеклянной губки (Porifera, Hexactinellida) в прибрежных водах Британской Колумбии, Канада. Мар. Экол. прог. сер. 283, 133–149. doi: 10.3354/meps283133
Полный текст CrossRef | Google Scholar
Лав, Г. Д., Грожан, Э., Сталвис, К., Фике, Д. А., Гротцингер, Дж. П., Брэдли, А. С., и др. (2009 г.). Ископаемые стероиды фиксируют появление демоспонгиев в криогенный период. Природа 457, 718–721. doi: 10.1038/nature07673
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Литл, Д. Э., и Валь, Э.Р. (2005). Реконструкции палеоокружающей среды с использованием современной аналоговой техники: влияние размера выборки и правил принятия решений. Голоцен 15, 554–566. doi: 10.1191/0959683605hl830rp
Полный текст CrossRef | Академия Google
Мальдонадо, М. (2009). Эмбриональное развитие демогубок-веронгид поддерживает независимое приобретение скелетов спонгинов в качестве альтернативы кремнистому скелету губок. биол. Дж. Линн. соц. 97, 427–447. doi: 10.1111/j.1095-8312.2009.01202.x
Полный текст CrossRef | Google Scholar
Мальдонадо, М. (2014). «Metazoans», в 90 524 Древе жизни. Систематика и эволюция живых организмов , редакторы В. Пабло и Р. Зардоя (Сандерленд, Массачусетс: Синауэр), 182–205.
Мальдонадо, М., Агилар, Р., Баннистер, Р. Дж., Белл, Дж. Дж., Конвей, К. В., Дейтон, П. К., и др. (2015а). «Губковые угодья как ключевые морские среды обитания: синтетический обзор типов, структуры, функциональных ролей и проблем сохранения», в Лесах морских животных: экология бентических биоразнообразия горячих точек , под редакцией С. Росси, Л. Браманти, А. Гори и К. Орехас Сако дель Валье (Cham: Springer International Publishing), 1–39.
Google Scholar
Мальдонадо М., Агилар Р., Бланко Дж., Гарсия С., Серрано А. и Пунсон А. (2015b). Агрегированные скопления литистидных губок: уникальная рифоподобная батиальная среда обитания с соответствующими палеонтологическими связями. PLoS ONE 10:e0125378. doi: 10.1371/journal.pone.0125378
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Maldonado, M., Carmona, M.C., Uriz, M.J., and Cruzado, A. (1999). Сокращение мезозойских рифообразующих губок объясняется дефицитом кремния. Природа 401, 785–788. doi: 10.1038/44560
CrossRef Полный текст | Google Scholar
Мальдонадо М., Кармона М. К., Веласкес З., Пуч А., Крузадо А., Лопес А. и др. (2005). Кремнистые губки как поглотитель кремния: упускаемый из виду аспект бентопелагического взаимодействия в морском цикле кремния. Лимнол. океаногр. 50, 799–809. doi: 10.4319/lo.2005.50.3.0799
Полный текст CrossRef | Google Scholar
Мальдонадо М., Наварро Л., Граса А., Гонсалес А. и Вакерисо И. (2011). Поглощение кремния губками: поворот к пониманию круговорота питательных веществ на континентальных окраинах. Науч. Респ. 1:30. doi: 10.1038/srep00030
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мальдонадо М., Рисго А., Буччи А. и Цлерб К. Р. (2010). Пересмотр баланса кремния на тропическом континентальном шельфе: постоянные запасы кремнезема в губках превосходят запасы диатомовых водорослей. Лимнол. океаногр. 55, 2001–2010 гг. doi: 10.4319/lo.2010.55.5.2001
CrossRef Полный текст | Google Scholar
Мальдонадо, М., и Янг, К.М. (1998). Пределы батиметрического распределения ороговевших губок: полевые испытания в глубокой воде. Мар. Экол. прог. сер. 174, 123–139. doi: 10.3354/meps174123
Полный текст CrossRef | Google Scholar
Малива, Р. Г., Нолл, А. Х., и Сивер, Р. (1989). Вековые изменения в распределении кремней: отражение эволюции биологического участия в круговороте кремнезема. Палеос 4, 519–532. doi: 10.2307/3514743
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Маррон А. О., Рэтклифф С., Уиллер Г. Л., Гольдштейн Р. Э., Кинг Н., Нот Ф. и др. (2016). Эволюция транспорта кремния у эукариот. Мол. биол. Эвол . 33, 3226–3248. doi: 10.1093/molbev/msw209
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Меркурио М., Коррьеро Г., Скалера Лиачи Л. и Гайно Э. (2000). Вариации содержания кремнезема и размеров спикул у Pellina semitubulosa ( Porifera: Demospongiae ). Мар биол. 137, 87–92. doi: 10.1007/s002270000336
Полный текст CrossRef | Google Scholar
Морроу, К., и Карденас, П. (2015). Предложение по пересмотренной классификации Demospongiae (Porifera). Фронт. Зоол . 12:7. doi: 10. 1186/s12983-015-0099-8
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мюллер, У. Э. Г., Краско, А., Ле Пеннек, Г., и Шредер, Х. К. (2003). Биохимия и клеточная биология образования кремнезема у губок. Микроск. Рез. Тех. 62, 368–377. doi: 10.1002/jemt.10402
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Павлик Дж. Р., МакМюррей С. Э., Эрвин П. и Зеа С. (2015). Нет доказательств ограниченности пищи карибских рифовых губок: ответ на slattery и более мелкие. Мар. Экол. прог. сер. 527, 281–284. doi: 10.3354/meps11308
Полный текст CrossRef | Google Scholar
Писера, А. (1997). Верхнеюрские кремнистые губки швабского Альба: таксономия и палеоэкология. Палеонтология Пол. 57, 1–216.
Писера, А. (2006). Палеонтология губок — обзор. Кан. Дж. Зул. 84, 242–261. doi: 10.1139/z05-169
Полный текст CrossRef | Google Scholar
Писера А. и Леви К. (2002). «Liithistid’ Demospongiae», в Systema Porifera. Руководство по классификации губок , редакторы JNA Hooper и RWM Van Soest (Нью-Йорк, штат Нью-Йорк: Kluwer Academic/Plenum Publishers), 299–301.
Google Scholar
Пшеславски, Р., Альварес, Б., Баттерсхилл, К., и Смит, Т. (2014). Биоразнообразие губок и экология возвышенности Ван-Димен и восточной части залива Джозефа Бонапарта, северная Австралия. Гидробиология 730, 1–16. doi: 10.1007/s10750-013-1799-8
CrossRef Full Text | Google Scholar
Пшеславски Р., Альварес Б., Кул Дж., Бридж Т., Кейли Дж. и Николс С. (2015). Влияние моделей биоразнообразия губок на управление морским заповедником в северной Австралии. PLoS ONE 10:e0141813. doi: 10.1371/journal.pone.0141813
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Przeslawski, R., Daniell, J., Anderson, T., Barrie, J. V., Heap, A., Hughes, M., et al. (2011). Места обитания на морском дне и опасности залива Жозефа Бонапарта и Тиморского моря, Северная Австралия . Geoscience Australia, Record 2011/40, 69.
Google Scholar
Racca, JMJ, Racca, R., Pienitz, R., and Prairie, YT (2007). PaleoNet: новое программное обеспечение для построения, оценки и применения передаточных функций на основе нейронных сетей в палеоэкологии. Дж. Палеолимнол. 38, 467–472. doi: 10.1007/s10933-006-9082-x
Полный текст CrossRef | Google Scholar
Raff, A.R., Marshall, C.R., and Turbeville, J.M. (1994). Использование последовательностей ДНК для раскрытия кембрийского излучения типов животных. год. Преподобный Экол. Сист. 25, 351–375. doi: 10.1146/annurev.es.25.110194.002031
CrossRef Full Text | Google Scholar
Рапп Х.Т., Януссен Д. и Тендал О.С. (2011). Известковые губки из абиссальных и батиальных глубин моря Уэдделла, Антарктида. Глубоководная служба спасения. II Топ. Стад. океаногр. 58, 58–67. doi: 10.1016/j.dsr2.2010.05.022
Полный текст CrossRef | Google Scholar
Рейнке, Т., и Бартель, Д. (1997). Кинетика поглощения кремнезема Halichondria panicea в Кильской бухте. Мар биол. 129, 591–593. doi: 10.1007/s002270050200
Полный текст CrossRef | Google Scholar
Риттербуш, К. А., Ботджер, Д. Дж., Корсетти, Ф. А., и Росас, С. (2014). Новые данные о роли кремнистых губок в экологии и развитии осадочных фаций восточной панталасса после триасово-юрского массового вымирания. Палеос 29, 652–668. doi: 10.2110/palo.2013.121
Полный текст CrossRef | Google Scholar
Риттербуш, К. А., Росас, С., Корсетти, Ф. А., Ботджер, Д. Дж., и Уэст, А. Дж. (2015). Андские губки обнаруживают долгосрочные сдвиги бентосной экосистемы после массового вымирания в конце триаса. Палеогеогр. Палеоклимат. Палеоэколь. 420, 193–209. doi: 10.1016/j.palaeo.2014.12.002
CrossRef Полный текст | Google Scholar
Рютцлер К. и Смит К. П. (1993). Род Terpios и новые виды в комплексе « Lobiceps» . Науч. 90 525 марта 57, 381–393.
Google Scholar
Сармьенто, Дж. Л., и Грубер, Н. (2006). Биогеохимическая динамика океана. Принстон, Нью-Джерси: Издательство Принстонского университета.
Google Scholar
Schönberg, CHL, and Barthel, D. (1997). Неорганический скелет демогубки Halichondria panicea . сезонность производства спикул в Балтийском море. 905:24 Мар. Биол. 130, 133–140. doi: 10.1007/s002270050232
Полный текст CrossRef | Google Scholar
Шредер Х.К., Перович-Оттштадт С., Ротенбергер М., Винс М., Швертнер Х., Батель Р. и др. (2004). Транспорт кремнезема в демогубке Suberites domuncula : эмиссионный анализ флуоресценции с использованием зонда PDMPO и клонирование потенциального переносчика. Биохим. J. 381 (часть 3), 665–673. doi: 10.1042/BJ20040463
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Сивер, Р. (1991). «Кремнезем в океанах: биолого-геохимическое взаимодействие», в 90 524 ученых на Гайе. Доклады, представленные на ежегодной конференции Чепмена Американского геофизического союза в марте 1988 г. , редакторы С. Х. Шнайдер и Дж. Б. Пенелопа (Кембридж, Массачусетс: MIT Press), 287–295.
Синка, К.Дж., и Аткинсон, Т.С. (1999). Метод взаимного климатического диапазона для реконструкции палеоклимата по растительным остаткам. Дж. Геол. соц. Лонд. 156, 381–396. doi: 10.1144/gsjgs.156.2.0381
Полнотекстовая перекрестная ссылка | Google Scholar
Слэттери М. и Лессер М. П. (2015). Трофическая экология губок от мелких до мезофотических глубин (от 3 до 150 м): комментарий к Pawlik et al. (2015). Мар. Экол. прог. сер. 527, 275–279. doi: 10.3354/meps11307
Полный текст CrossRef | Google Scholar
Spalding, MD, Fox, H.E., Allen, G.R., Davidson, N., Ferdaña, Z.A., Finlayson, M., et al. (2007). Морские экорегионы мира: биорайонирование прибрежных и шельфовых районов. Биологические науки 57, 573–583. doi: 10.1641/B570707
Полный текст CrossRef | Google Scholar
Stone, AR (1970). Сезонные изменения размера спикул у hymeniacidon perleve. J. Mar. Biol. доц. Великобритания 50, 343–348. doi: 10.1017/S0025315400004562
CrossRef Полный текст | Google Scholar
Телфорд, Р. Дж., и Биркс, Х. Дж. Б. (2005). Секретное предположение передаточных функций: проблемы с пространственной автокорреляцией при оценке производительности модели. Q. Наука. Ред. 24, 2173–2179. doi: 10.1016/j.quascirev.2005.05.001
CrossRef Полный текст | Google Scholar
Уриз, М. Дж., Турон, X., Бесерро, М., и Агелл, Г. (2003). Кремнистые спикулы и каркасы скелета губок: происхождение, разнообразие, ультраструктурные закономерности и биологические функции. Микроск. Рез. Тех. 62, 279–299. doi: 10.1002/jemt.10395
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Vacelet, J. (1988). Indications de deeperur données par les Spongiaires dans les milieux benthiques actuels. Géologie Méditerranéenne 15, 13–26.
Google Scholar
Валисано Л., Поццолини М., Джовине М. и Серрано К. (2012). Отложение биокремнезема в морской губке Petrosia ficiformis (Poiret, 1789): модель примморфов выявляет зависимость спикулогенеза от времени. Hydrobiologia 687, 259–273. doi: 10.1007/s10750-011-0987-7
CrossRef Полный текст | Google Scholar
Van Soest, RWM (1994). «Схемы распространения Demosponge», в S ponges in Time and Space , редакторы RWM Van Soest, TMG Van Kempen и JC Braekman (Роттердам: Balkema), 213–223.
Google Scholar
Van Soest, R.W.M., Boury-Esnault, N., Hooper, J., Rützler, K., de Voogd, N.J., Alvarez, B., et al. (2017). Всемирная база данных Porifera . Доступно в Интернете по адресу: http://www.marinespecies.org/porifera. Последняя консультация (по состоянию на 23 марта 2017 г.).
Van Soest, R.W.M., Boury-Esnault, N., Vacelet, J., Dohrmann, M., Erpenbeck, D., De Voogd, N.J., et al. (2012). Глобальное разнообразие губок (Porifera). PLoS ONE 7:e35105. doi: 10.1371/journal.pone.0035105
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Wang X., Schloßmacher U., Wiens M., Batel R., Schröder HC и Müller WEG (2012a). Силикатеины, силикатеиновые интеракторы и клеточное взаимодействие в скелетогенезе губок: образование спикул, подобных стекловолокну. FEBS J. 279, 1721–1736. doi: 10.1111/j.1742-4658.2012.08533.x
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Wang, X., Schroder, H.C., Wang, K., Kaandorp, J.A., and Muller, W.E.G. (2012b). Генетические, биологические и структурные иерархии при формировании спикул губок: от мягких золь-гелей до твердых трехмерных композитных структур из кремнезема. Мягкая материя 8, 9501–9518. doi: 10.1039/c2sm25889g
CrossRef Полный текст | Google Scholar
Wang, X., Wiens, M., Schröder, H.C., Jochum, K.P., Schloßmacher, U., Götz, H., et al. (2011а). Окружной рост спикул за счет перицеллюлярного отложения кремнезема у гексактинеллидной губки Monorhaphis chuni. Дж. Эксп. биол. 214, 2047–2056. doi: 10.1242/jeb.056275
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Wang X., Wiens M., Schröder H.C., Schloßmacher U., Pisignano D., Jochum K.P. и др. (2011б). Эвагинация клеток контролирует образование и созревание биокремнезема во время образования спикул у губок. PLoS ONE 6:e20523. doi: 10.1371/journal.pone.0020523
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Уитни Ф., Конвей К., Томсон Р., Барри В., Крауттер М. и Мунгов Г. (2005). Океанографическая среда обитания губчатых рифов на континентальном шельфе Западной Канады. Прод. Полка Рез. 25, 211–226. doi: 10.1016/j.csr.2004.09.003
Полный текст CrossRef | Google Scholar
Вилле М., Саттон Дж., Эллвуд М. Дж., Сэмбридж М., Махер В., Эггинс С. и др. (2010). Фракционирование изотопов кремния в морских губках: новая модель для понимания вариаций изотопов кремния в губках. Планета Земля. науч. лат. 292, 281–289. doi: 10.1016/j.epsl.2010.01.036
Полный текст CrossRef | Google Scholar
Сяо С., Ху Дж., Юань X., Петрушка Р. Л. и Цао Р. (2005). Сочлененные губки из формации хетанг нижнего кембрия на юге Аньхоя, Южный Китай: их возраст и значение для ранней эволюции губок. Палеогеогр. Палеоклимат. Палеоэколь. 220, 89–117. doi: 10.1016/j.palaeo.2002.02.001
Полный текст CrossRef | Google Scholar
Yin, Z., Zhu, M., Davidson, E.H., Bottjer, D.J., Zhao, F., and Tafforeau, P. (2015). Окаменелость тела губчатого типа с клеточным разрешением, датируемая 60 млн лет до кембрия. Проц. Натл. акад. науч. США 112, E1453–E1460. doi: 10.1073/pnas.1414577112
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Юрасовски, К., и Расмонт, Р. (1984). Дифференцировка склероцитов у пресноводных губок, выращенных на среде с низким содержанием кремнезема. Дифференциация 25, 5–9. doi: 10.1111/j.1432-0436. 1984.tb01330.x
Полный текст CrossRef | Google Scholar
Зеа, С. (1987). Колумбийский Эспонхас-дель-Карибе. Bogota: Editorial Catálogo Científico, 285.
Google Scholar
Губка Определение и значение | Dictionary.com
- Основные определения
- Синонимы
- Викторина
- Связанный контент
- Примеры
- Британский
- Медицинский
- Научный
- Идиомы и фразы
- тесто, поднятое на дрожжах, особенно перед замесом, как для хлеба.
- легкий сладкий пудинг пористой консистенции, приготовленный из желатина, яиц, фруктового сока или другого вкусоароматического материала и т. д.
Очищайте сковороду сразу после каждого использования, пока она еще теплая, вручную, используя горячую воду и губку.
Советы от Элоизы: санитарная обработка и дезинфекция — не одно и то же|Элоиза Элоиза|29 октября 2020 г.|Washington Post
Затем я наношу базовый слой краски с помощью губки для макияжа — в краску добавлен латекс чтобы обеспечить гибкость и избежать растрескивания — по всей маске.
Создание масок для Хэллоуина, которые не отстой|Юджин Робинсон|16 октября 2020 г.|Ozy
Этот дизайн отлично подходит для тех, кто привык пользоваться губкой, и он по-прежнему достаточно прочный, чтобы чистить даже самые твердые поверхности, такие как гриль после приготовления пищи или чугун после вяления рыбы.
Металлические скрубберы для поддержания вашей чугунной сковороды в первозданном виде|Компания PopSci Commerce|23 сентября 2020 г.|Popular-Science
Представьте себе технологию в виде гигантской губки, вставленной чуть ниже уровня земли.
Норвежский стартап превращает засушливые пустыни в плодородные пахотные земли|Ванесса Бейтс Рамирес|19 августа 2020 г.
|Singularity Hub Они используют морские губки, чтобы защитить свои клювы, пока выкапывают пищу на морском дне.
Дельфины могут научиться у своих сверстников тому, как использовать раковины в качестве инструментов|Джек Дж. Ли|6 августа 2020 г.|Новости науки для студентов
Свифт надеется, что роль жертвы, дерзкой губки «ненависти», продается.
Почему мы должны ненавидеть «Хейтеры возненавидят»|Тим Тиман|25 августа 2014|DAILY BEAST
Никакого майонеза, только масло, которое, как бисквит, впиталось в булочку.
Мой большой маслянистый ролл с лобстером: мы пришли, мы захватили, мы победили|Скотт Биксби|7 июня 2014 г.|DAILY BEAST
Санта-Клаус угощает рисовым пудингом в Дании, бисквитом в Чили, кулкулами в Индии и пирогами с фаршем в Великобритании.
Что общего у губки в форме смайлика, приспособления для приучения кошек к туалету и зонта без помощи рук?
«Танк с акулами»: Роберт Херявек рассказывает о деньгах, Марке Кьюбане и гоночных автомобилях|Анна Классен|4 октября 2013 г.
|DAILY BEASTЧто еще более важно, его крестовый поход против весла из губки спас его любимый вид спорта от непредсказуемой судьбы.
Марти Рейзман: Волшебный мошенник, спасший классическую игру|Гарольд Эванс|10 декабря 2012|DAILY BEAST
Аристид посетил мадам Кокеро, которая развлекла его сладким вином Фронтиньян, сухими бисквитами и беседой.
Веселые приключения Аристида Пужоля|Уильям Дж. Локк
Там стоял сосуд, полный уксуса: поэтому они положили губку, полную уксуса, на иссоп и поднесли его ко рту.
Его последняя неделя|Уильям Э. Бартон
Однажды ей показалось, что шляпа ее папы выглядит грубой и ржавой: так что же она сделала, кроме как помыла ее губкой, смоченной в воде.
Детская, январь 1873 г., том. XIII.|Разное
После того, как оно окажется на бутылке, возьмите немного лучшего сладкого масла и чистой губкой тщательно промокните кружево до самых сокровенных складок.
Книга женского этикета и руководство по вежливости|Флоренс Хартли
Завершите их, окунув губку в размер, приготовленный путем кипячения изингласса в воде, и протрите изнаночную сторону.
Книга женского этикета и Руководство по вежливости | Флоренс Хартли , обычно встречающийся в сложных сидячих колониях, в которых пористое тело поддерживается волокнистым, известковым или кремнистым каркасом скелета
кусок легкого пористого эластичного каркаса некоторых губок с высокой впитывающей способностью, используемых для купания, уборки и т. д. См. также губку
любой из ряда легких пористых эластичных материалов, напоминающих губку
другое слово для губки (по определению 1)
неформальный человек, злоупотребляющий алкоголем
дрожжевое тесто, особенно перед замесом0003
пористый металл, полученный электролизом или восстановлением соединения металла без плавления или спекания и способный поглощать большое количество газоплатиновой губки
протирание губкой
вбрасывание губки См.
вбрасывание (по определению 4)глагол
(tr; часто следует off или down) очищать (что-либо) путем протирания или протирания влажной или мокрой губкой путем протирания влажной или мокрой губкой или тканью
(когда tr, часто следует вверх) впитывать (жидкости, особенно когда пролито) как губка
(tr часто фолл мимо off) получать (что-то) от (кого-то), полагаясь на его щедростьк губке еда от кого-то
(ввод; часто следует от случая к случаю) для получения средств к существованию, благосостояния и т. д., неоправданно (от) он обтирает губками своих друзей
(ввод) собирать губки
См. также губку
Производные формы губки
губчатый, прилагательное
Происхождение слова для губки
Старый английский, от латинского spongia, от греческого
Английский словарь Коллинза — полное и полное цифровое издание 2012 г.
© William Collins Sons & Co. Ltd., 1979, 1986 © HarperCollins
Publishers 1998, 2000, 2003, 2005, 2006, 2007, 2009, 2012.Любое из многочисленных водных беспозвоночных животных типа Porifera.
Легкий, волокнистый, впитывающий скелет некоторых из этих организмов.
Кусок абсорбирующего пористого материала, такого как целлюлоза, пластик или резина, используемый специально для стирки и чистки.
Марлевая подушечка, используемая для впитывания крови и других жидкостей, например, в хирургии или при перевязке ран.
Противозачаточная губка.
v.
Смывать, увлажнять или впитывать губкой.
Медицинский словарь Стедмана The American Heritage®
Авторское право © 2002, 2001, 1995 от компании Houghton Mifflin. Опубликовано компанией Houghton Mifflin.Научное определение губки
губка
[ spŭnj ]
Любое из многочисленных водных, главным образом морских беспозвоночных животных типа Porifera. Для губок характерен пористый скелет, обычно содержащий сложную систему каналов, состоящую из волокнистого материала, кремнистых или известковых спикул.
Вода, проходя через поры, приносит в организм пищу. Губки обитают на всех глубинах моря, ведут сидячий образ жизни и часто образуют колонии неправильной формы, прикрепленные к подводной поверхности. Губки считаются самыми примитивными представителями животного мира, поскольку у них отсутствует нервная система и дифференцированные ткани или органы тела. Взрослые особи не имеют движущихся частей, а личинки свободно плавают. Губки обладают большими регенеративными способностями, причем некоторые виды способны регенерировать полный взрослый организм из фрагментов размером всего в одну клетку. Губки впервые появились в раннем кембрийском периоде и, возможно, произошли от простейших. Также называется порифераном. См. примечание при регенерации.Легкий, волокнистый, гибкий, впитывающий скелет некоторых из этих организмов, используемый для купания, уборки и других целей.
Кусок пористого пластика, резины, целлюлозы или другого материала, сходного по впитывающей способности с этим каркасом и используемого для тех же целей.
Научный словарь American Heritage®
Copyright © 2011. Опубликовано издательством Houghton Mifflin Harcourt Publishing Company. Все права защищены.Другие идиомы и фразы с губкой
губка
В дополнение к идиоме, начинающейся со слова губка
- губка на
, см. также:
- вброс в губку
Словарь идиом американского наследия®
Авторские права © 2002, 2001, 1995, издательство Houghton Mifflin Harcourt Publishing Company. Опубликовано издательством Houghton Mifflin Harcourt Publishing Company.300-летние арктические губки лакомятся трупами своих разлагающихся вымерших соседей
Губкам в среднем 300 лет, а многим даже больше. Они вмещают сложное сообщество микроорганизмов в симбиотических отношениях, что способствует здоровью и питанию губок.
(Изображение предоставлено: Alfred-Wegener-Institut/PS101 AWI OFOS system/Antje Boetius, [email protected])На подводной горе в Северном Ледовитом океане живет сообщество губок с омерзительным секретом.
Имея мало пищи в бедной питательными веществами воде, губки выживают, переваривая останки давно умерших животных, которые когда-то населяли пики подводных гор, где сейчас живут губки. И веками они пировали на трупах своих вымерших соседей.Ученые недавно обнаружили этих жутких существ на хребте Лангсет, части бывшей вулканической подводной горы в Центральной Арктике, на глубине от 1640 до 1,969 футов (от 500 до 600 метров), где температура колеблется чуть выше нуля. В этих ледяных глубинах исследователи обнаружили тысячи губок на площади 15 квадратных километров.
В некоторых частях глубоководного океана, где не хватает питательных веществ, экосистемы морского дна часто группируются вокруг гидротермальных жерл (также называемых выходами), которые обеспечивают тепло и пищу, но вулканическая активность в этой части подводной горы прекратилась тысячи лет назад. Нет и сильных океанских течений, которые могли бы нести пищу губкам сверху или снизу. Но губки обнаружили богатый и обильный запас пищи: растворенные соединения с кладбища давно вымерших трубчатых червей и двустворчатых моллюсков, которые губки переваривают с небольшой помощью симбиотика 9.
1583 бактерии .Связанные: 10 самых странных мест на Земле, где есть жизнь сообщество; они также собрали образцы губок и окружающей их среды с помощью дистанционно управляемого водолазного робота под названием «Нереида подо льдом», согласно исследованию, опубликованному 8 февраля в журнале 9.1583 Связь с природой .
Мало того, что на пиках подводных гор скопились тысячи губок, многие из них стали довольно большими, достигая 3 футов (1 м) в диаметре, говорит ведущий автор исследования Тереза Морганти, научный сотрудник Института Макса Планка. по морской микробиологии в Бремене, Германия. И многие из губок активно размножались, показывая «существенное почкование», — написали ученые в исследовании.
«Главный вопрос заключался в том, как такое сообщество могло выжить в этой области?», — сказал Морганти Live Science. «Мы предположили, что они могли использовать местный источник пищи — в данном случае остатки этого древнего сообщества сипов».
Ученые обнаружили удивительно богатую и густонаселенную экосистему на вершинах потухших подводных вулканов в арктических глубинах. Среди них преобладали губки, растущие там в большом количестве и достигающие внушительных размеров. (Изображение предоставлено: Alfred-Wegener-Institut/PS101 AWI OFOS system/Antje Boetius, [email protected])
(открывается в новой вкладке)
Под губками исследователи обнаружили плотную биомассу, состоящую в основном из трубок, оставленных после морские черви, которые вымерли, когда вулканическая активность подводной горы прекратилась примерно 2000–3000 лет назад, и следа губки на ископаемых матах показали, где губки кормились, прежде чем осесть на сохранившихся останках. Многим отдельным губкам было не менее 300 лет, и в них обитали разнообразные микроорганизмы. Авторы исследования сообщили, что бактерии типа Chloroflexi, вероятно, сыграли важную роль в деградации окаменелых трубчатых червей и высвобождении растворенного органического вещества, которое обеспечивало сытость губок.
Но если губки уже веками обитают в этом месте, а сейчас их тысячи, то не грозит ли им истощение запасов пищи? По словам Морганти, вероятно, нет, так как у губок исключительно медленный метаболизм.
ДОПОЛНИТЕЛЬНОЕ СОДЕРЖАНИЕ
«Поскольку это крупные особи, живущие в центральной части Арктики, где температура низкая, их метаболизм в целом очень низок», — объяснила она. «Они не потребляют этот источник пищи очень быстро, поэтому я думаю, что у них там много еды».
Тем не менее, даже если неограниченный выбор ископаемых трупов губок не будет исчерпан в ближайшее время, изменение климата может создать более непосредственную угрозу выживанию губок. Поверхность моря над хребтом Лангсет обычно покрыта слоем льда. Но по мере того, как Арктика нагревается и поверхностный морской лед тает, все больше питательных веществ будет дрейфовать с поверхности моря на расположенные ниже подводные горы. По словам Морганти, если пища станет более обильной в глубинах океана, другие морские виды могут начать двигаться и нарушить глубоководную среду обитания губок.
2 уровень сложности.
[spuhnj]
/ spʌndʒ /
Сохрани это слово!
См. синонимы для: губка / губка / похожая на губку на Thesaurus.com
Показывает уровень оценки в зависимости от сложности слова.
сущ.
любое водное, главным образом морское животное типа Porifera, имеющее пористую структуру и обычно роговой, кремнистый или известняковый внутренний скелет или каркас, встречающееся большими сидячими колониями.
легкий, податливый, пористый, волокнистый скелет или каркас некоторых животных или колоний этой группы, особенно представителей родов Spongia и Hippospongia, из которых удалены живые вещества, характеризующийся тем, что они легко впитывают воду и становятся мягкими при намокании во время сохранение прочности: используется при купании, при протирании или чистке поверхностей и т. д.
любые другие подобные вещества, часто пористая резина или целлюлоза, используемые для стирки или чистки.
губка для ванн.
человек или предмет, который свободно впитывает что-то: Его разум — губка, собирающая исторические данные.
лицо, которое постоянно занимает у других или живет за счет других; тунеядец; паразит.
Сленг: пренебрежительный и оскорбительный. заядлый алкоголик, который часто находится в состоянии алкогольного опьянения.
Металлургия. пористая масса металлических частиц, таких как платина, полученная восстановлением оксида или очищенного соединения при температуре ниже точки плавления.
Хирургия. стерильная хирургическая повязка из абсорбирующего материала, обычно хлопчатобумажной марли, для вытирания или поглощения гноя, крови или других жидкостей во время хирургической операции.
Кулинария.
одноразовый кусок пенополиуретана, пропитанный спермицидом, для введения во влагалище в качестве противозачаточного средства.
глагол (используется с дополнением), губкой, губкой.
протирать или протирать влажной губкой, а также увлажнять или очищать.
для удаления влажной губкой или влажной губкой (обычно с последующим удалением и т. д.).
для стирания или стирания с помощью губки или как с помощью губки (часто с последующим удалением).
впитывать или впитывать с помощью губки или как с помощью губки (часто следует вверх): набирать воду губкой.
одолжить, использовать или получить, навязывая чужому добродушию, дружбе, гостеприимству и т. п.: Он выпросил 40 баксов у своего друга и отправился в город.
Керамика. украшать (керамический предмет), протирая его губкой, смоченной краской.
глагол (используется без дополнения), губка, губка.
для приема или всасывания жидкости путем абсорбции.
для сбора губок.
жить за счет других (часто за этим следует включение или выключение): он вернулся домой и какое-то время протирал свою семью.
ДРУГИЕ СЛОВА ДЛЯ губки
6 пиявка.
12 стирк.
См. синонимы слова губка на Thesaurus.com
ВИКТОРИНА
Сыграем ли мы «ДОЛЖЕН» ПРОТИВ. «ДОЛЖЕН» ВЫЗОВ?
Следует ли вам пройти этот тест на «должен» или «должен»? Это должно оказаться быстрым вызовом!
Вопрос 1 из 6
Какая форма используется для указания обязательства или обязанности кого-либо?
Идиомы про губку
кинуть в губку, Неформал. признать поражение; урожай; сдаться: результаты досрочных выборов были сильно против него, но он не был готов бросить губку.
Происхождение губки
Впервые отмечено до 1000 года; Среднеанглийское существительное губка, губка, губка, древнеанглийское существительное губка, губка, от латинского spongia, Spongea, от греческого spongiā́; глагол от существительного
ДРУГИЕ СЛОВА СЛОВО губка
губка·без губки, прилагательныегубчатая, прилагательнаягубчатая, наречие· губчатая, прилагательное губка-мешок, губка для ванны, бисквит, губка, губка-пух
Dictionary.com Полный текст
На основе Random House Unabridged Dictionary, © Random House, Inc. 2022
Слова, относящиеся к губке
паразит, пьяница, пьяница, рамми, типплер, алкаш, бездельник, бездельник, нахлебник, прихлебатель, пиявка, попрошайка , scrounger, beg, cadge, chisel, freeload, hustle, panhandle