Содержание
Новый вид цианобактерий проливает свет на эволюцию кислородного фотосинтеза
Из растущего в Панаме мха выделен новый вид цианобактерий, названный Anthocerotibacter panamensis. На сегодняшний день у A. panamensis самое простое строение и самый простой фотосинтетический аппарат среди известных цианобактерий. Благодаря этому ученые смогли многое понять про историю появления кислородного фотосинтеза, преобразившего нашу планету. В частности, теперь мы знаем, что у общего предка цианобактерий еще не было тилакоидов (структур, внутри которых происходит фотосинтез), а также представляем себе минимальный набор белков, необходимых для кислородного фотосинтеза.
Монстры, преобразившие мир
Представьте, что на Земле появился новый вид бактерий, который в качестве побочного продукта выделяет ядовитый газ хлор. Из-за них в короткий период времени атмосфера наполняется хлором. Небо из голубого становится желто-зеленым. Сначала гибнут люди и другие позвоночные, позже, по мере роста концентрации токсичного газа, Земля лишается растений и насекомых. Лишь некоторые бактерии успевают приспособиться к ядовитому газу и научиться использовать его для получения энергии. Через миллионы лет они дадут начало новым формам жизни на Земле. Это выглядит как апокалиптическая сцена из фильма-катастрофы, но похожее событие на самом деле произошло на Земле около двух с половиной миллиардов лет назад. За исключением того факта, что жизнь на тот момент была представлена только бактериями и археями — эукариоты еще не успели возникнуть.
Лишь некоторые бактерии успевают приспособиться к ядовитому газу и научиться использовать его для получения энергии. Через миллионы лет они дадут начало новым формам жизни на Земле. Это выглядит как апокалиптическая сцена из фильма-катастрофы, но похожее событие на самом деле произошло на Земле около двух с половиной миллиардов лет назад. За исключением того факта, что жизнь на тот момент была представлена только бактериями и археями — эукариоты еще не успели возникнуть.
Около двух с половиной миллиардов лет назад появились цианобактерии — группа бактерий, которая научилась проводить фотосинтез, используя в качестве донора электронов обычную воду (напомню, что как при дыхании, так и при фотосинтезе поток электронов от одного белкового комплекса к другому непосредственно дает энергию для перекачки протонов и создания протонного градиента). К тому времени фотосинтез уже давно существовал, но он требовал неорганического «сырья» типа сероводорода, которое еще надо найти. Цианобактерии в этом плане оказались в явном выигрыше: воды вокруг было в буквальном смысле залейся, солнечного света и углекислого газа тоже хватало, так что необходимые им ресурсы оказались практически ничем не ограниченными.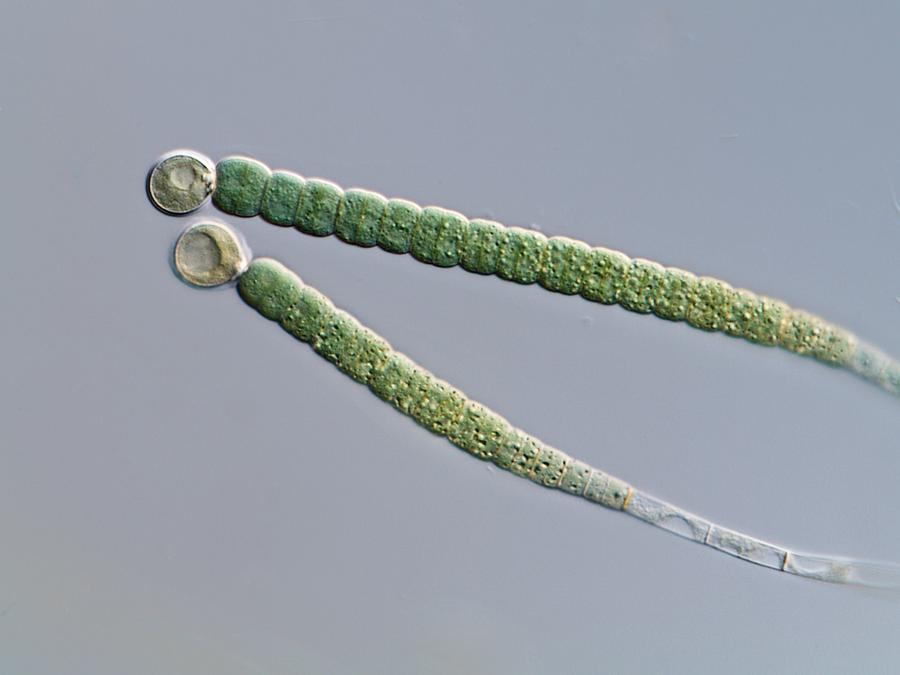 Благодаря этому их численность резко подскочила. Но у цианобактерий был один важный недостаток: в процессе такого выгодного фотосинтеза в качестве отхода выделялся токсичный кислород, получающийся при расщеплении воды.
Благодаря этому их численность резко подскочила. Но у цианобактерий был один важный недостаток: в процессе такого выгодного фотосинтеза в качестве отхода выделялся токсичный кислород, получающийся при расщеплении воды.
Если бы злой рок забросил вас примерно на 2,9 миллиарда лет назад, то вы бы задохнулись. В тогдашней атмосфере почти не было кислорода. В ней преобладали углекислый газ, азот и метан — в общем, все то, что за предыдущие полтора миллиарда лет успели произвести вулканы. С химической точки зрения такая атмосфера создавала восстановительную среду, то есть в целом была склонна отдавать электроны, а не отнимать их. Как ни странно, это оказалось на руку зарождающейся жизни: вся органика очень уязвима к окислителям, и восстановительная атмосфера была поистине комфортной колыбелью для жизни. Это была оптимальная среда для первых обитателей Земли.
Массивное выделение кислорода цианобактериями произвело такой же эффект, как если бы сейчас атмосфера наполнилась хлором. В ряду электроотрицательности неметаллов кислород на втором месте, а хлор — только на четвертом. Поэтому кислород оказывал на тогдашнюю жизнь такой же негативный эффект, как хлор — на современную. Токсичность обоих газов заключается в их высочайшей окислительной способности, к которой чрезвычайно чувствительна вся земная жизнь: она построена из органических макромолекул, легко повреждающихся сильными окислителями.
Поэтому кислород оказывал на тогдашнюю жизнь такой же негативный эффект, как хлор — на современную. Токсичность обоих газов заключается в их высочайшей окислительной способности, к которой чрезвычайно чувствительна вся земная жизнь: она построена из органических макромолекул, легко повреждающихся сильными окислителями.
Мы сейчас не замечаем пагубного влияния кислорода лишь потому, что эволюционно приспособились к нему. Точнее, к нему приспособились наши предки. Помимо того, что мы утилизируем его при дыхании, мы вооружены множеством ферментов и антиоксидантов для нейтрализации активных форм кислорода. Наши клетки не замечают этого яда лишь потому, что постоянно «сражаются» с ним и используют в своих целях. В общем, цианобактерии своим кислородом полностью преобразили биосферу и метаболизм многих живых существ, совершив настоящую «кислородную революцию».
Помимо цианобактерий, оксигенный фотосинтез могут осуществлять растения и многие группы водорослей. Но все они осуществляют его при помощи пластид, которые происходят либо от поглощенных когда-то цианобактерий, либо. .. от других фотосинтезирующих эукариот, имевших пластиды, произошедшие от цианобактерий. Как пишет Александр Марков в книге «Рождение сложности»: «Некоторые биологи говорят, пользуясь метафорическим языком, что растения — всего лишь удобные «домики» для проживания цианобактерий. По сути дела цианобактерии не только изобрели кислородный фотосинтез, но и по сей день сохранили за собой «эксклюзивные права» на его осуществление».
.. от других фотосинтезирующих эукариот, имевших пластиды, произошедшие от цианобактерий. Как пишет Александр Марков в книге «Рождение сложности»: «Некоторые биологи говорят, пользуясь метафорическим языком, что растения — всего лишь удобные «домики» для проживания цианобактерий. По сути дела цианобактерии не только изобрели кислородный фотосинтез, но и по сей день сохранили за собой «эксклюзивные права» на его осуществление».
Если представить, какую роль растения и водоросли играют в современном облике нашей планеты, то станет ясно, что мы живем в мире, сформированном цианобактериями.
Цианобактерии «придумали» не только фотосинтез. Если посмотреть на цианобактерии под микроскопом, в глаза бросится еще одно их отличие: среди них много многоклеточных видов, существующих в виде нитей из многих клеток (среди эукариот аналогами такой жизненной формы являются нитчатые водоросли). Причем речь идет именно о настоящей многоклеточности: клетки в составе нити обмениваются метаболитами и сигнальными молекулами. А у наиболее «продвинутых» цианобактерий (например, представителей рода Nostoc) существует дифференцировка на разные типы клеток: одни клетки (вегетативные) фотосинтезируют, а другие (гетероцисты) занимаются фиксацией азота.
А у наиболее «продвинутых» цианобактерий (например, представителей рода Nostoc) существует дифференцировка на разные типы клеток: одни клетки (вегетативные) фотосинтезируют, а другие (гетероцисты) занимаются фиксацией азота.
Молекулярно-филогенетические исследования показывают, что многоклеточность у цианобактерий — настолько же древнее явление, как и сам кислородный фотосинтез (B. E. Schirrmeister et al., 2013. Evolution of multicellularity coincided with increased diversification of cyanobacteria and the Great Oxidation Event). Это подтверждают и окаменелости, удивительно похожие на современные формы многоклеточных цианобактерий (см. статью Сергея Ястребова Кислородная революция и Земля-снежок). Можно уверенно утверждать, что цианобактерии точно находятся в ряду первых «изобретателей» многоклеточности. Более того, есть предположение, что какими-то молекулярными «находками» для поддержания межклеточной кооперации они поделились с нами. Есть гипотеза, что именно они изобрели каспазы — белки запрограммированной клеточной гибели, — и далее все остальные многоклеточные организмы позаимствовали эти белки у них (N. Lane, 2008. Marine microbiology: Origins of Death). Также не исключено, что у цианобактерий мы «научились» синтезировать оксилипины — универсальные молекулы межклеточной сигнализации (см. Зачем бактериям и простейшим липоксигеназы?, «Элементы», 31.10.2020). Впрочем, это пока только гипотеза.
Lane, 2008. Marine microbiology: Origins of Death). Также не исключено, что у цианобактерий мы «научились» синтезировать оксилипины — универсальные молекулы межклеточной сигнализации (см. Зачем бактериям и простейшим липоксигеназы?, «Элементы», 31.10.2020). Впрочем, это пока только гипотеза.
Цианобактериальные инновации
Такой власти над миром цианобактериям удалось добиться лишь потому, что они научились при фотосинтезе расщеплять воду вместо сероводорода. А сделать это не так просто — вновь из-за того, что кислород является сильным окислителем, и высвободить его из воды труднее, чем серу из сероводорода (чем сильнее окислитель, тем труднее он сам окисляется). Чтобы выпустить в атмосферу такой химически активный газ (отняв у него электроны), одной фотосистемы (электрон-транспортной цепи фотосинтеза) недостаточно. История цианобактерий началась с того, что в результате горизонтального переноса генов они смогли объединить в себе фотосистемы двух «образцов», существовавшие у пурпурных бактерий и зеленых серобактерий. Вместе они развивают гораздо больший оксилительно-восстановительный потенциал, чем по отдельности, что и позволяет отрывать электроны от воды. В целом можно сказать, что объединение двух фотосистем позволило цианобактериям высвобождать в атмосферу мощный окислитель. И это же привело к катастрофе в биосфере.
Вместе они развивают гораздо больший оксилительно-восстановительный потенциал, чем по отдельности, что и позволяет отрывать электроны от воды. В целом можно сказать, что объединение двух фотосистем позволило цианобактериям высвобождать в атмосферу мощный окислитель. И это же привело к катастрофе в биосфере.
Большинство известных на сегодняшний день цианобактерий имеют весьма совершенный фотосинтетический аппарат — не только с точки зрения биохимии, но и с точки зрения структуры. Цепь переноса электронов, задействованная в фотосинтезе, очень похожа на дыхательную цепь, но в митохондриях и хлоропластах (а также у аэробных бактерий и большинства цианобактерий) эти цепи располагаются в разных местах. У митохондрий (рис. 3) они встроены во внутреннюю мембрану, которая происходит от цитоплазматической мембраны когда-то поглощенного нашими далекими предками бактериального симбионта. А у бактерий дыхательная цепь находится в самой цитоплазматической мембране. Цепи переноса электронов выкачивают протоны на внешнюю сторону мембраны, наполняя ими межмембранное пространство. Получившийся градиент концентрации протонов в буквальном смысле вращает АТФ-синтазу, «чеканящую» универсальную энергетическую валюту клетки — АТФ.
Получившийся градиент концентрации протонов в буквальном смысле вращает АТФ-синтазу, «чеканящую» универсальную энергетическую валюту клетки — АТФ.
А в хлоропластах и у большинства цианобактерий имеется замкнутая система мембранных дисков и пластинок, отделенных от внутренней мембраны — тилакоидов. Именно на мембранах тилакоидов размещаются электрон-транспортные цепи фотосинтеза, использующие энергию солнечного света для закачивания протонов внутрь тилакоидов. Их плоская форма оптимальна для создания минимального объема внутри при максимальной поверхности, позволяющей разместить максимальное число электрон-транспортных комплексов. Такая конструкция концентрирует протоны куда более эффективно, чем внутренняя мембрана митохондрий, позволяя синтезировать АТФ куда более эффективно, чем это происходит при дыхании.
Помимо эффективного механизма концентрации протонов, современные цианобактерии и растения имеют мощные молекулярные «антенны», улавливающие свет и передающие его молекулам хлорофилла. У растений это светособирающие комплексы, а у цианобактерий — фикобилисомы (рис. 4).
У растений это светособирающие комплексы, а у цианобактерий — фикобилисомы (рис. 4).
Белки фикобилисомы содержат в качестве небелковой части фикобилины (похожие на наш билирубин), которые непосредственно «собирают» кванты света. Аналогичные вещества используют растения — только у них светособирающий комплекс устроен по-другому.
Чтобы эффективно использовать свет, цианобактериям пришлось обзавестись и другой инновацией. Как известно, у многих многоклеточных организмов — как растений, так и у животных — независимо развились сенсоры дня и ночи и регуляция ритмов активности в зависимости от времени суток, называемых циркадными (циркадианными) ритмами.
У растений за это отвечают криптохромы — особые белки, позволяющие растению чувствовать свет. Хоть растения и не способны видеть, но свет от тьмы они «отличают». У животных, в том числе у человека, криптохромы тоже задействованы в регуляции циркадных ритмов (они находятся в ядрах гипоталамуса, регулирующих цикл сна и бодрствования).

Большинству бактерий такие механизмы без надобности, но цианобактериям они жизненно необходимы: их биохимия целиком и полностью зависит от солнечного света. Поэтому у них есть внутренние «биологические часы», подсказывающие, когда какие реакции проводить. Они состоят из трех белков (не родственных криптохромам), вступающих в циклические биохимические реакции и тем самым образующих своеобразный «молекулярный маятник» (хотя в хронобиологии подобные системы называются осцилляторами). Что интересно, эту систему можно вживить в любую другую бактерию — и она тоже будет осуществлять те или иные биохимические функции «по часам» (см. В бактерию без циркадных ритмов встроили «часы» от цианобактерии, «Элементы», 15.06.2015).
Наличие «биологических часов» тоже позволяет большинству известных цианобактерий эффективно использовать солнечный свет. В общем, у цианобактерий есть все, чтобы в «промышленных» масштабах разлагать воду и производить сильный окислитель — кислород. Возникает вопрос: а как они пришли к такому состоянию? Как способность к фотосинтезу развивалась в ходе эволюции?
Древо жизни цианобактерий
Ближайших «родственников» цианобактерий мы можем обнаружить. .. у себя в кишечнике, а также в пресных водоемах. Они называются мелайнабактериями (Melainabacteria), что в буквальном переводе с греческого означает «черные бактерии» (в противоположность цианобактериям — «лазурным бактериям»). Это название весьма говорящее: никто из мелайнабактерий не умеет осуществлять фотосинтез. Оксигенный фотосинтез, очевидно, появился после разделения цианобактерий и мелайнабактерий. Ясности в вопросе, как развился оксигенный фотосинтез, это не прибавляет.
.. у себя в кишечнике, а также в пресных водоемах. Они называются мелайнабактериями (Melainabacteria), что в буквальном переводе с греческого означает «черные бактерии» (в противоположность цианобактериям — «лазурным бактериям»). Это название весьма говорящее: никто из мелайнабактерий не умеет осуществлять фотосинтез. Оксигенный фотосинтез, очевидно, появился после разделения цианобактерий и мелайнабактерий. Ясности в вопросе, как развился оксигенный фотосинтез, это не прибавляет.
Но сами цианобактерии — не однородная группа. На ее филогенетическом дереве выделяется базальная (то есть отходящая раньше всего) ветвь — так называемые глеобактерии (Gloeobacteria, рис. 5). Все остальные цианобактерии выделяются в группу Phycobacteria, но чаще всего в англоязычной литературе встречается наименование «кроновые цианобактерии» (crown cyanobacteria) — из-за того, что именно они образуют «крону» филогенетического дерева цианобактерий.
В изучении эволюции цианобактерий глеобактерии представляют собой такую же ценность, как ланцетники при изучении эволюции хордовых (см. Функциональный анализ генома ланцетника проливает новый свет на эволюцию позвоночных, «Элементы», 26.11.2018). Это наиболее рано обособившиеся представители, по которым легче всего попытаться реконструировать эволюционные изменения, приведшие к появлению целой новой группы. Глеобактерии на данный момент — практически единственное доступное нам звено, переходное от нефотосинтезирующих мелайнабактерий к мастерам высокоэффективного кислородного фотосинтеза. И практически единственный наш шанс ответить на вопрос: как появились эти монстры, испортившие земную атмосферу 2,5 млрд лет назад?
Функциональный анализ генома ланцетника проливает новый свет на эволюцию позвоночных, «Элементы», 26.11.2018). Это наиболее рано обособившиеся представители, по которым легче всего попытаться реконструировать эволюционные изменения, приведшие к появлению целой новой группы. Глеобактерии на данный момент — практически единственное доступное нам звено, переходное от нефотосинтезирующих мелайнабактерий к мастерам высокоэффективного кислородного фотосинтеза. И практически единственный наш шанс ответить на вопрос: как появились эти монстры, испортившие земную атмосферу 2,5 млрд лет назад?
Проблема в том, что доступных данных по глеобактериям до настоящего времени было мало — всего два вида были выращены в культуре и описаны экспериментально: Gloeobacter violaceus и Gloeobacter kilaueensis. G. violaceus стал основным модельным организмом для изучения эволюции оксигенного фотосинтеза. На поверку он оказался устроен гораздо проще «кроновых» цианобактерий. Во-первых, он одноклеточный и не имеет циркадных ритмов. Во-вторых, у него отсутствуют тилакоиды. Если фотосистемы «кроновых» цианобактерий и фотоавтотрофных эукариот «закачивают» протоны в люмен (просвет) тилакоидов, то у G. violaceus они просто выкачиваются на внешнюю сторону цитоплазматической мембраны, в межмембранное пространство (см. рис. 3). В итоге способом создания протонного градиента G. violaceus больше всего напоминает не «кроновые» цианобактерии и хлоропласты, а наши митохондрии и их альфа-протеобактериальных предшественников — у них протонный градиент генерируется так же.
Во-вторых, у него отсутствуют тилакоиды. Если фотосистемы «кроновых» цианобактерий и фотоавтотрофных эукариот «закачивают» протоны в люмен (просвет) тилакоидов, то у G. violaceus они просто выкачиваются на внешнюю сторону цитоплазматической мембраны, в межмембранное пространство (см. рис. 3). В итоге способом создания протонного градиента G. violaceus больше всего напоминает не «кроновые» цианобактерии и хлоропласты, а наши митохондрии и их альфа-протеобактериальных предшественников — у них протонный градиент генерируется так же.
Таким образом, общий план строения клетки G. violaceus очевидно проще, чем у «кроновых» цианобактерий. Помимо этого, у G. violaceus фикобилисомы устроены проще, чем у других описанных цианобактерий (об этом позже) и отсутствуют некоторые компоненты фотосистем. Что, казалось бы, интуитивно ясно — можно ожидать, что рано «ответвившийся» представитель цианобактерий будет более примитивным.
Есть у него и еще одно биохимическое отличие, связанное с биосинтезом каротиноидов. Каротиноиды — это те самые желтые, красные и оранжевые пигменты, которые расширяют спектральный диапазон поглощаемого света и помогают более эффективно использовать его энергию при фотосинтезе. Но каротиноиды умеют синтезировать не только растения и цианобактерии. Они также синтезируются многими нефотосинтезирующими бактериями и грибами. Но при этом в природе существует два способа синтезировать каротиноиды — два варианта биохимического пути их синтеза (рис. 6). Различия между этими путями касаются одного «участка» — введения в молекулу фитоена (phytoene) дополнительных двойных связей, что превращает его в ликопин. В «растительном» пути, используемом растениями и большинством цианобактерий, это превращение осуществляется в три стадии тремя разными ферментами-десатуразами. А в «бактериальном» пути, используемом грибами и нефотосинтезирующими бактериями, это превращение протекает в одну стадию и катализируется одной десатуразой.
Каротиноиды — это те самые желтые, красные и оранжевые пигменты, которые расширяют спектральный диапазон поглощаемого света и помогают более эффективно использовать его энергию при фотосинтезе. Но каротиноиды умеют синтезировать не только растения и цианобактерии. Они также синтезируются многими нефотосинтезирующими бактериями и грибами. Но при этом в природе существует два способа синтезировать каротиноиды — два варианта биохимического пути их синтеза (рис. 6). Различия между этими путями касаются одного «участка» — введения в молекулу фитоена (phytoene) дополнительных двойных связей, что превращает его в ликопин. В «растительном» пути, используемом растениями и большинством цианобактерий, это превращение осуществляется в три стадии тремя разными ферментами-десатуразами. А в «бактериальном» пути, используемом грибами и нефотосинтезирующими бактериями, это превращение протекает в одну стадию и катализируется одной десатуразой.
Как оказалось, G. violaceus использует «бактериальный» тип биосинтеза каротиноидов, соответствующий нефотосинтезирующим бактериям. Возникает интуитивный соблазн тоже отнести эту особенность к проявлениям «примитивизма».
Возникает интуитивный соблазн тоже отнести эту особенность к проявлениям «примитивизма».
Но выводы о «примитивном» или «изначальном» состоянии в данном случае являлись бы очень спекулятивными и поспешными. В самом деле, какие-то из перечисленных особенностей могли развиться у G. violaсeus уже после его обособления от других цианобактерий, — то есть он мог претерпеть что-то вроде «эволюционной дегенерации» (если говорить совсем грубо). Тем самым не исключено, что общий предок всех цианобактерий уже имел и тилакоиды, и биосинтез каротиноидов «растительного» типа, а глеобактер просто их утратил. Двух видов было абсолютно недостаточно, чтобы делать далеко идущие эволюционные выводы. Требовалось выделить и охарактеризовать кого-нибудь еще из глеобактерий — и желательно не из рода Gloeobacter.
Откуда не ждали
Новый вид был открыт неожиданно. Открыла его группа под руководством Фэй-Вэй Ли (Fai-Wei Li), сотрудника Института Бойса Томпсона (входящего в Корнеллский университет), которая занимается выделением цианобактериальных симбионтов из антоцеротовых мхов. У любого из этих мхов есть какой-нибудь бактериальный симбионт из числа многоклеточных цианобактерий, осуществляющий для мха фиксацию атмосферного азота (так же, как клубеньковые бактерии снабжают фиксированным азотом растения семейства Бобовые). Этих-то симбионтов и искали исследователи. И очень удивились, когда из мха Leiosporoceros dussii (рис. 7) вместо нитчатой цианобактерии выделилась одноклеточная глеобактерия. Это была редкая удача, но тем не менее она была неожиданной.
У любого из этих мхов есть какой-нибудь бактериальный симбионт из числа многоклеточных цианобактерий, осуществляющий для мха фиксацию атмосферного азота (так же, как клубеньковые бактерии снабжают фиксированным азотом растения семейства Бобовые). Этих-то симбионтов и искали исследователи. И очень удивились, когда из мха Leiosporoceros dussii (рис. 7) вместо нитчатой цианобактерии выделилась одноклеточная глеобактерия. Это была редкая удача, но тем не менее она была неожиданной.
Это было тем более неожиданно, что новый вид, названный Anthocerotibacter panamensis (рис. 1 и рис. 8; буквально это название означает «бактерия антоцеротового мха из Панамы») никак не годился на роль азотфиксирующего: у него нет гена, необходимого для фермента, отвечающего за фиксацию азота. Такого симбионта мху держать незачем, так что непонятно, как он оказался в его талломе. Авторы выдвинули два предположения: или Anthocerotibacter panamensis случайно занял «пустующее» место постоянного симбионта (который, скорее всего, существует, просто не был обнаружен в этом образце), или только он выдержал процедуру поверхностной стерилизации, применяемую авторами для удаления бактерий, попавших из почвы.
Еще одним неожиданным моментом стало то, что Anthocerotibacter panamensis оказался ближайшим родственником не выделенных пока глеобактерий, обитающих в Арктике и филогенетически входящих в так называемую «полярную кладу». Это тоже было неожиданно — сам-то A. panamensis происходит из тропиков (а конкретно — из Панамы, о чем и говорит его название). Возможно, что «полярная клада» вовсе не такая уж полярная, а возможно, что новый вид действительно просто экзотический.
Говоря «не выделенных пока глеобактерий», я имел в виду, что их пока никто не видел под микроскопом и не вырастил в культуре. А откуда мы тогда знаем, что бактерии «полярной клады» существуют? Это известно из метагеномного исследования (C. L. Grettenberger et al., 2020. A phylogenetically novel cyanobacterium most closely related to Gloeobacter). В ходе таких исследований секвенируется вся ДНК, найденная в каком-нибудь образце почвы или воды. Секвенирование и последующий биоинформатический анализ могут с некоторой степенью надежности воссоздать геномы еще не выделенных в культуре бактерий, оказавшихся в образце.
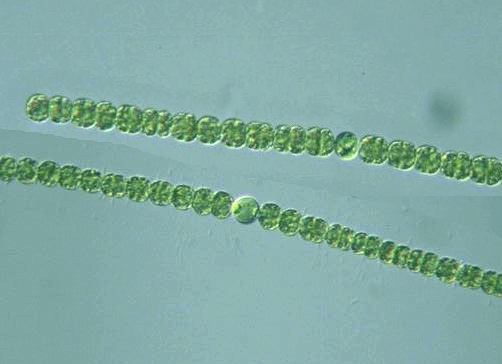
Новый вид оказался во многом похож на глеобактера. Он также одноклеточный, не имеет циркадных ритмов и тилакоидов. Это надежно указывает на то, что многоклеточность, «биологические часы» и тилакоиды появились у общего предка «кроновых» цианобактерий, а общий предок всех цианобактерий их еще не имел. Очевидно, он при фотосинтезе использовал ту же менее эффективную схему, когда протонный градиент создается на мембране клетки, а способ повысить напряжение за счет тилакоидов был более поздним приобретением. «Теперь мы можем быть почти уверены, что тилакоиды развились у фикобактерий» — говорит Фэй-Вэй Ли. Как и следовало ожидать, в плане отсутствия тилакоидов глеобактерии представляют собой сохранившийся реликт, самое примитивное состояние. Но так произошло не со всеми признаками.
С биосинтезом каротиноидов все получилось совсем наоборот, чем можно было бы предполагать. Оказалось, что у A. panamensis каротиноиды синтезируются по «растительному» пути, как и у наиболее «продвинутых» цианобактерий. Это позволяет предположить, что этот путь сформировался уже у общего предка всех цианобактерий — и лишь у некоторых глеобактерий, включая G. violaceus, он потом заменился на «бактериальный». Такой пример вторичного «упрощения», если так можно выразиться.
Это позволяет предположить, что этот путь сформировался уже у общего предка всех цианобактерий — и лишь у некоторых глеобактерий, включая G. violaceus, он потом заменился на «бактериальный». Такой пример вторичного «упрощения», если так можно выразиться.
Фотосинтезирует, но плохо
Но самые интересные находки касались фотосинтеза. У нового вида фотосинтетический аппарат оказался самым примитивным из известных на сегодняшний день. Прежде всего, у него самые простые фикобилисомы. Вспомним, что у «кроновых» цианобактерий и некоторых фотосинтезирующих эукариот фикобилисомы имеют вид антенны-локатора или вентилятора (рис. 9, сверху). Основание антенны образовано белком аллофикоцианином (АФЦ), а «лепестки» построены из фикоцианина (ФЦ) и фикоэритрина (ФЭ). Иногда место последнего занимает фикоэритроцианин (ФЭЦ). Но у G. violaceus фикобилисома проще и имеет вид стопки из белков, или, если изображать упрощенно, столбика (рис. 9, в середине). Таким образом, если сравнивать фикобилисомы с техническими сооружениями, то у «кроновых» цианобактерий имеется тарелка локатора, а у глеобактера — что-то вроде телевизионных антенн, которые можно встретить на крышах домов. Но состоит антенна из тех же белков: АФЦ, ФЦ и ФЭ.
Но состоит антенна из тех же белков: АФЦ, ФЦ и ФЭ.
Структура фикобилисомы A. panamensis пока экспериментально не определена, так что говорить о ней (и пытаться ее нарисовать!) можно лишь предположительно. Одно мы знаем точно — она проще, чем даже у глеобактера: в ней отсутствует фикоэритрин. Таким образом, фикобилисома антроцеротибактера пока что претендует на звание самой простой и примитивной по белковому составу. Внизу на рис. 9 я нарисовал, как это могло бы выглядеть, если форма фикобилисомы антоцеротибактера такая же, как у глеобактера. Продолжая наши аналогии, получившуюся конструкцию можно сравнить с комнатной телевизионной антенной, — настолько она упрощена. Но, повторюсь, формы фикобилисомы глеобактера мы пока не знаем. И это лишь делает более интригующим вопрос: как выглядит самая простая фикобилисома на свете?
Впрочем, у нового вида не только фикобилисомы устроены максимально просто. Сами фотосистемы — «рабочий механизм» фотосинтеза — у него тоже упрощены (рис. 10). В фотосистеме I отсутствует треть всех белков, из которых строится та же фотосистема у «кроновых» цианобактерий. Правда, у глеобактера фотосистема I устроена, по всей видимости, так же. А вот по части фотосистемы II антоцеротибактер пока удерживает безусловный рекорд простоты: у него отсутствует пять из 23 белков (у G. violaceus нет четырех белков, у G. kilaueensis — трех). Подобная примитивность заставляет биохимиков еще раз задуматься над вопросом, каков минимальный «набор» белков, необходимых для оксигенного фотосинтеза? Это представляет большой интерес для синтетической биологии.
10). В фотосистеме I отсутствует треть всех белков, из которых строится та же фотосистема у «кроновых» цианобактерий. Правда, у глеобактера фотосистема I устроена, по всей видимости, так же. А вот по части фотосистемы II антоцеротибактер пока удерживает безусловный рекорд простоты: у него отсутствует пять из 23 белков (у G. violaceus нет четырех белков, у G. kilaueensis — трех). Подобная примитивность заставляет биохимиков еще раз задуматься над вопросом, каков минимальный «набор» белков, необходимых для оксигенного фотосинтеза? Это представляет большой интерес для синтетической биологии.
Не менее интересным для исследователей оказался вопрос: а насколько эффективный фотосинтез способна производить бактерия с таким примитивным фотосинтетическим аппаратом? Для этого ученые протестировали скорость роста нового вида в отсутствие органических питательных веществ и при наличии одного лишь света, а также определили скорость выработки кислорода. Оказалось, что, «питаясь» одним светом, новая цианобактерия растет очень медленно: почти трое суток ей требуется на одно деление, даже если свет яркий. При более тусклом свете она делится и вовсе раз в неделю. Это гораздо медленнее, чем у «кроновых» цианобактерий (их представитель Synechocystis sp. PCC 6803 делится каждые 8 часов) и сопоставимо с одним из рекордсменов медленного роста совсем из другой «оперы» — микобактерией туберкулеза. Это показывает, насколько неэффективен фотосинтетической аппарат антоцеротибактера. Но при этом скорость выделения кислорода была лишь примерно вдвое ниже, чем у «кроновой» цианобактерии Synechocystis sp. PCC 6803. То есть, если бы антроцеротибактер 2,5 млрд лет назад «работал» в одиночку, он все же имел бы шансы произвести кислородную революцию и преобразить мир. Только, может быть, на полмиллиарда лет позже.
При более тусклом свете она делится и вовсе раз в неделю. Это гораздо медленнее, чем у «кроновых» цианобактерий (их представитель Synechocystis sp. PCC 6803 делится каждые 8 часов) и сопоставимо с одним из рекордсменов медленного роста совсем из другой «оперы» — микобактерией туберкулеза. Это показывает, насколько неэффективен фотосинтетической аппарат антоцеротибактера. Но при этом скорость выделения кислорода была лишь примерно вдвое ниже, чем у «кроновой» цианобактерии Synechocystis sp. PCC 6803. То есть, если бы антроцеротибактер 2,5 млрд лет назад «работал» в одиночку, он все же имел бы шансы произвести кислородную революцию и преобразить мир. Только, может быть, на полмиллиарда лет позже.
Рисунки созданы с помощью biorender.com.
Источник: Nasim Rahmatpour, Duncan A. Hauser, Jessica M. Nelson, Pa Yu Chen, Juan Carlos Villarreal A., Ming-Yang Ho, Fay-Wei Li. A novel thylakoid-less isolate fills a billion-year gap in the evolution of Cyanobacteria // Current Biology. 2021. DOI: 10.1016/j.cub.2021.04.042.
2021. DOI: 10.1016/j.cub.2021.04.042.
Георгий Куракин
Цианобактерии убивают нервные клетки с помощью фальшивой аминокислоты
Наука
close
100%
Эффективные лекарства против нейродегенеративных заболеваний могут быть найдены с помощью сине-зеленых водорослей, которые синтезируют токсичную аминокислоту, встраивающуюся в белки, — эта трансформация приводит к гибели двигательных нейронов.
Сине-зеленые водоросли – цианобактерии – древнейшая группа живых организмов, за миллиарды лет эволюции приспособившаяся к самым разнообразным условиям обитания на планете. Способность превращать углекислый газ и азот воздуха в необходимые для роста и размножения вещества позволяет им селиться в самых, казалось бы, безжизненных местах, например на голых скалах. Выживают они даже в хлорированной водопроводной воде.
Болезнь Альцгеймера можно лечить заранее
Накануне Всемирного дня борьбы с болезнью Альцгеймера ученые опубликовали описание нового молекулярного. ..
..
20 сентября 15:09
Попадая в благоприятные условия – богатые органикой водоемы, цианобактерии достаточно быстро размножаются, заполняя собой пространство и вызывая знакомое многим явление, получившее название «цветение воды». Есть среди них полезные и даже съедобные виды, например спирулина. Есть и ядовитые.
Ядовитые синтезируют нейротоксин – аминокислоту β-N-метил-амино-50-аланин (BMAA).
Эта аминокислота не участвует в формировании белковых молекул, но при попадании в организм человека способна выводить из строя двигательные нейроны, нанося необратимые повреждения центральной нервной системе.
Болезнь двигательных нейронов (боковой амиотрофический склероз, известный также как болезнь Лу Герига или болезнь Шарко) – неизлечимое терминальное нейродегенеративное заболевание, поражающее людей в возрасте 40–60 лет. Заболеваемость в мире составляет два случая в год на 100 тыс. человек. Болезнь убивает двигательные нейроны головного и спинного мозга и постепенно приводит к параличу, не затрагивая интеллект больного. Более чем в 90% случаев причины заболевания неизвестны и способы лечения неясны.
Среди людей, страдающих этой болезнью, известный физик Стивен Хокинг – один из немногих, чей диагноз был подтвержден, а состояние стабилизировано.
Ученые из Австралии попытались выяснить, что именно происходит, когда токсичная аминокислота цианобактерий попадает в организм человека. Для Австралии эта проблема достаточно актуальна, поскольку водные артерии страны часто подвергаются агрессивному нашествию сине-зеленых водорослей. Наиболее памятным было «цветение воды», распространившееся летом 1991/1992 года на расстояние более 1000 километров.
Сто возбудителей менингита
В связи со вспышкой менингита в Ростовской области «Газета.Ru» рассказывает о том, что это за заболевание, как…
18 июня 16:20
Взаимосвязь между повреждением двигательных нейронов и употреблением пищи или воды, загрязненной цианобактериями, была установлена достаточно давно, в 70-е годы прошлого века, но механизм действия токсина оставался неясным.
Впервые BMAA была обнаружена в 1972 году в организме населяющих остров Гуам представителей коренной народности чаморро, которые, как выяснилось, в 100 раз чаще заболевают болезнью Шарко, чем другие жители планеты. Чаморро используют в пищу саго – крахмал саговой пальмы. Его добывают из сердцевины стволов растений. В сыром виде саго ядовито из-за высокого содержания BMAA. Токсин обезвреживают специальной обработкой. Другой пищевой традицией этой народности является употребление в пищу летучих мышей, которые также питаются саговниками. То есть изрядную долю BMAA чаморро получают вместе с обычной для них пищей.
С тех пор исследователи стали обращать более пристальное внимание на вспышки болезни двигательных нейронов среди людей, живущих вблизи «цветущих» водоемов, в том числе среди тех, кто употреблял в пищу моллюсков, накопивших токсические количества цианобактерий, а также среди солдат НАТО, воевавших в Персидском заливе в 1990–1991 годах.
Исследователи из Технологического университета Сиднея во главе с Кеном Роджерсом, в содружестве с ботаником Полом Коксом и учеными из Института народной медицины Вайоминга (США) обнаружили, что
токсичная BMAA имитирует в организме человека другую аминокислоту – серин, встраиваясь вместо нее в синтезируемые нашим организмом обычные белки.
Нейролептики иссушают мозг
Прием нейролептиков — основного лекарственного средства, которое назначается больным шизофренией…
18 сентября 14:07
Такие ошибочные белки со временем накапливаются и влекут за собой гибель клеток. Результаты работы опубликованы в журнале PLOS ONE.
«Общим свойством всех нейродегенеративных заболеваний является то, что в нервных клетках начинают накапливаться определенные белки, причем в огромных количествах, – пояснила Рэйчел Данлоп, первый автор статьи. – Когда перегруженность клетки этими белками достигает критических значений, она совершает суицид, самоубийство. В нашем исследовании мы показали, что BMAA может служить механизмом, запускающим этот процесс».
Для того чтобы изучить механизм действия токсичной аминокислоты, австралийские ученые использовали в качестве модельной системы линии клеток человека.
Как оказалось, достаточное количество L-серина, правильной аминокислоты, с которой за место в белковой молекуле и конкурирует BMAA, может предотвратить выработку неправильного белка организмом и снизить вероятность развития болезни.
Авторы исследования считают, что L-серин может стать новым эффективным лекарством против данной группы нейродегенеративных заболеваний.
Подписывайтесь на «Газету.Ru» в Новостях, Дзен и Telegram.
Чтобы сообщить об ошибке, выделите текст и нажмите Ctrl+Enter
Новости
Дзен
Telegram
Картина дня
«Интерфакс»: военный Су-34 упал на жилой дом в Ейске
Военная операция РФ на Украине. День 236-й
Онлайн-трансляция военной спецоперации РФ на Украине — 236-й день
Москва завершает мобилизацию. Уклонистов обещают не искать, а повестки аннулировать
Собянин объявил о завершении частичной мобилизации в Москве
В Госдуме предложили внести в законы термин «добровольческие формирования»
Mash: в России больше не будет оригинальной «Виагры»
Мобилизованных из Забайкальского края отправили домой из-за проблем с алкоголем
Депутат Хинштейн заявил, что в «Лолите» Набокова нет пропаганды педофилии
Новости и материалы
Худрук «Модерна» Грымов назвал преимущества театра перед кино
Очевидец рассказал, что Су-34 уже горел перед падением в Ейске
В Минобороны РФ назвали причину падения самолета в Ейске
Психологи выяснили, что птичье пение избавляет от паранойи и тревожности
Судья «Ледникового периода» перепутал Милохина с актрисой Дарьей Мороз
Команда гражданской миссии Евросоюза прибыла в Армению
Bloomberg: ежегодный ущерб от дефицита рабочих рук обойдется Германии в $85 млрд
«Милохин, держи!»: судья «Ледникового периода» испугалась за Медведеву
Опубликовано фото с места столкновения «КамАЗа» и поезда в Тамбовской области
Лолита станет ведущей третьего сезона шоу «Суперстар»
Худрук «Модерна» Грымов признался, что разочаровался в отечественном кино
Алферова: в номере Медведевой и Милохина не было такого спектакля, как у Арзамасовой и Кацалапова
Боец UFC Махачев рассказал, как начал заниматься единоборствами
RT: по неподтвержденным данным, у самолета, упавшего в Ейске, отказал двигатель
Тарасова: я сама сделалась чудесной после выступления Медведевой и Милохина
Кокорин: всем будет лучше, чтобы я не появлялся в сборной России
Военный самолет упал на жилой дом в Ейске
Звезда «Ангелов Чарли» Дрю Бэрримор заявила, что шесть лет не занималась сексом
Все новости
Женщин на моряков. Пушилин анонсировал новый обмен пленными с Украиной
Пушилин заявил об обмене пленными с Украиной по формуле 110 на 110
Ружье против дронов: российские войска начали применять «Гарпун-3»
«Ступор», «Гарпун», «Алабуга»: как электромагнитное оружие влияет на ход спецоперации
«Сотни населенных пунктов обесточены». К чему привели новые удары по Украине
Мэр Владимир Кличко заявил о нескольких взрывах в Шевченковском районе в центре Киева
«Дума обязана работать». Не всем депутатам понравилась идея отмены их брони от мобилизации
В Госдуму внесли законопроект об отмене отсрочки от призыва для депутатов и сенаторов
«Петр I под знаменем Христа служил Антихристу»: интервью с худруком театра «Модерн»
Худрук театра «Модерн» Грымов обвинил американские компании в разрушении российского проката
Новая специальность: где и как мобилизованных учат управлять дронами
В Минобороны рассказали о подготовке пилотов-операторов БПЛА
Король хип-хопа: Эминему — 50
«Сербия должна присоединиться к санкциям». ЕС принуждает Белград отвернуться от России
ЕС принуждает Белград отвернуться от России
«Ъ»: Евросоюз усилил давление на Сербию и требует ввести антироссийские санкции
«Аня, твой Паша зарегистрировался на сайте знакомств»: мама сыновей с аутизмом – о предательстве мужа
Личная история матери о том, как ее муж сбежал от сыновей с аутизмом
От обвинений в гомосексуальности до семейного счастья: как сегодня живет 50-летний Таркан
Как сегодня выглядит и чем занимается 50-летний Таркан
«Годовой эксперимент SIRIUS по моделированию полета на Луну состоится в любых условиях»
Руководитель проекта SIRIUS назвал цели годового эксперимента по моделированию полета на Луну
«Нет сомнений, на какой стороне должен стоять Израиль». В Тель-Авиве призвали поставить оружие Киеву
В Тель-Авиве призвали поставить оружие Киеву
Министр по делам диаспоры Израиля выступил за оказание военной помощи Украине
США пообещали одинаковый ответ на ядерный удар любого масштаба по Украине
Салливан заявил, что ответ США на ядерный удар по Украине не будет зависеть от масштаба
Вода Magazine — Цианобактерии (сине-зеленые водоросли) и токсины цианобактерий: риски, инструменты контроля и технологии устранения
Немногочисленные отечественные исследования по проблематике цианотоксинов посвящены, как правило, определению концентраций цианотоксинов в водах поверхностных источников питьевого водоснабжения. Данные по обнаружению цианотоксинов в питьевой воде редки и статистически непредставительны. С учетом этого представляет интерес переводной обзор трех наиболее значимых цитотоксинов, встречающихся в водах почти всех регионов США и перечисленных в третьем Списке загрязнителей-кандидатов (CCL) — микроцистина-LR, анатоксина-а и цилиндроспермопсина.
 В обзоре рассматриваются аспекты воздействия этих цитотоксинов на здоровье человека, инструменты аналитического контроля и эффективность различных технологий водоподготовки в устранении или инактивации указанных цитотоксинов.
В обзоре рассматриваются аспекты воздействия этих цитотоксинов на здоровье человека, инструменты аналитического контроля и эффективность различных технологий водоподготовки в устранении или инактивации указанных цитотоксинов.
Сразу надо отметить, что другие цианотоксины, такие как сакситоксины и анатоксины (S), также встречаются в водах США, но в общем рассматриваются как намного менее обычные (немного реже встречающиеся). Поэтому настоящий обзор не касается этих хорошо известных токсинов цианобактерий, таких как паралитический токсин моллюсков (семейство сакситоксинов), анатоксин-а(S), лингбиатоксины, или загрязнителей запаха и вкуса, причиной которых являются цианобактерии.
Фоновая информация
Закон о безопасной питьевой воде (SDWA) защищает общественное здоровье путем нормирования национальных общественных запасов питьевой воды и их источников: рек, озер, водохранилищ, родников и скважин подземных вод. SDWA обязывает Агентство охраны окружающей среды США (US EPA) публиковать список ненормируемых загрязнителей, которые встречаются или, как ожидается, могут встречаться в общественных системах водоснабжения в США и могут представлять риск в питьевой воде. Данный список известен как Список загрязнителей-кандидатов (CCL). Для получения более полной информации о программе CCL следуйте по ссылке: http://water.epa.gov/scitech/ drinkingwater/dws/ccl/.
Данный список известен как Список загрязнителей-кандидатов (CCL). Для получения более полной информации о программе CCL следуйте по ссылке: http://water.epa.gov/scitech/ drinkingwater/dws/ccl/.
Цианотоксины, включенные в последний CCL, продуцируются несколькими видами цианобактерий (цианобактерии известны как сине-зеленые водоросли). Наиболее распространенные цианотоксины — это токсины пептидной природы класса микроцистинов. Существует по меньшей мере 80 известных микроцистинов, включая микроцистин-LR, который считается одним из самых токсичных. Более десятка стран, включая Канаду, Бразилию, Новую Зеландию и Австралию, располагают разработанными нормативами или руководствами по содержанию микроцистинов в питьевой воде и рекреационных водах. Большинство руководств по питьевой воде основывается на предлагаемом Всемирной организацией здравоохранения (ВОЗ) уровне содержания микроцистина-LR в питьевой воде в 1,0 мкг/л. В настоящее время в США не существует федеральных нормативных руководств по цианобактериям или их токсинам в питьевой воде и рекреационных водах.
Причины вредного водорослевого цветения цианобактерий
Цианобактерии — это фотосинтезирующие бактерии, имеющие ряд общих свойств с водорослями и естественно встречающиеся в озерах, реках, прудах и прочих поверхностных водах. Как и другие типы водорослей, цианобактерии могут при благоприятных условиях быстро размножаться в воде, вызывая «цветение». Ниже описаны условия, усиливающие рост цианобактерий. Некоторые типы цианобактерий, например, Anabaena flos1 aquae, имеют газоносные полости, позволяющие им плавать на поверхности или под поверхностью, в зависимости от условий освещения или уровней концентраций питательных веществ. Это служит причиной концентрации цианобактерий на поверхности воды, вызывая типичный цвет горохового супа или сине-зеленую пену. Некоторые цианобактерии, подобно Planktothrix agardhii, встречаются в донных отложениях и всплывают в результате шторма или иных взмучиваний отложений. Другие цианобактерии при массовом развитии могут оставаться взвешенными в столбе воды (Cylindrospermopsis sp. ), вызывая всеобщее изменение цветности воды.
), вызывая всеобщее изменение цветности воды.
Факторы, влияющие на развитие и продолжительность цветения цианобактерий, охватывают интенсивность освещения и общую продолжительность освещения солнцем, доступность питательных веществ (особенно фосфора), температуру воды, рН, увеличение количества осадков, потока воды (является ли водный массив непроточным или быстро текущим), стабильность водного столба. Хотя условия развития цветения в большей части США являются наиболее благоприятными в поздней части лета, взаимодействия всех этих факторов вызывают существенные сезонные и годичные колебания численности цианобактерий. Некоторые продуцирующие токсины штаммы могут встречаться в начале лета, в то время как другие — только во второй половине летнего периода.
Цветение цианобактерий может быть вредным для окружающей среды, животных и здоровья человека. При разложении наросшей при цветении биомассы потребляется кислород, создавая условия гипоксии — недостатка кислорода, приводящие к отмиранию животных и растений.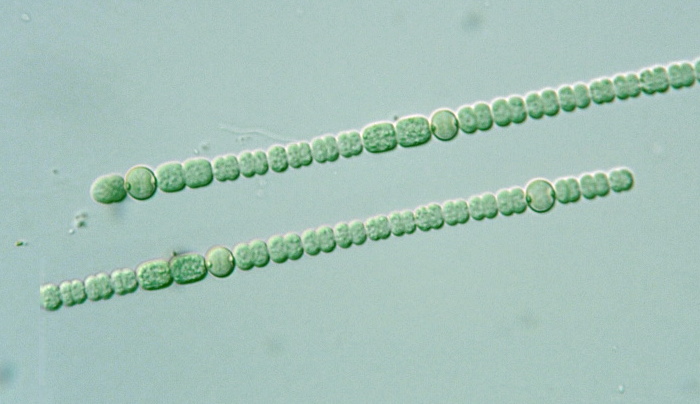 При благоприятных условиях освещенности и обеспечения питательными веществами некоторые виды цианобактерий продуцируют вторичные метаболиты, известные как цианотоксины. Распространенные токсинобразующие цианобактерии перечислены в таблице 1.
При благоприятных условиях освещенности и обеспечения питательными веществами некоторые виды цианобактерий продуцируют вторичные метаболиты, известные как цианотоксины. Распространенные токсинобразующие цианобактерии перечислены в таблице 1.
Условия, вызывающие образование цианобактериями цианотоксинов, не поняты до конца. Некоторые виды, способные к образованию токсинов, могут не продуцировать токсины при всех условиях среды. Данные виды часто входят во множество обычных формирующих цветение родов. Существуют как нетоксичные, так и токсичные разновидности большинства обычных токсинпродуцирующих цианобактерий, поэтому невозможно сказать, является ли вид токсичным или нетоксичным при его определении. Даже при наличии токсинпродуцирующих цианобактерий в действительности они могут не образовывать токсины. Более того, некоторые виды цианобактерий могут продуцировать многочисленные типы и разновидности цианотоксинов. Доступны молекулярно биологические тесты, способные определить, несет ли цианобактерия, например, Microcystis, ген токсина; для того, чтобы определить, действительно ли цианобактерия продуцирует токсин, необходим количественный анализ по определению цианотоксинов. При этом загрязненная цианобактериями вода может встречаться без каких-либо проблем привкуса и запаха.
При этом загрязненная цианобактериями вода может встречаться без каких-либо проблем привкуса и запаха.
В большинстве случаев токсины цианобактерий существуют внутриклеточно в цитоплазме и остаются внутри клетки. Разновидности анатоксина-а и микроцистина обнаруживались внутриклеточно приблизительно в течение 95% времени ростовой стадии цветения. Для данных видов токсины высвобождаются в воду при отмирании или разрушении клеток, при разрыве клеточной мембраны (экстрацеллюлярные внеклеточные токсины). Однако у других видов, например, в случае цилиндроспермопсина, значительное количество токсина может выделяться естественным образом в воду живыми клетками цианобактерий; по литературным данным соотношение примерно 50% внутриклеточных к 50% внеклеточным токсинам. Внеклеточные токсины могут абсорбироваться на глине и органических взвесях в водном столбе и в целом более сложно удаляются, чем внутриклеточные токсины.
Влияние цианоксинов на здоровье
Цианоксины включают нейротоксины (воздействующие на нервную систему), гепатотоксины (воздействующие на печень) и дерматотоксины (воздействующие на кожу). Присутствие высоких уровней цианотоксинов в рекреационных и питьевых водах может вызывать у человека широкий спектр симптомов (таблица 1), включая повышение температуры, головную боль, мышечные и суставные боли, кожные волдыри, желудочные спазмы, понос, рвоту, язвы в ротовой полости и аллергические реакции. Данные эффекты могут проявляться в течение нескольких минут или дней после воздействия. В некоторых случаях могут происходить приступы, печеночная недостаточность, остановка дыхания и (редко) смерть. Существуют доказательства, что долгосрочное воздействие микроцистинов и цилиндроспермопсина может способствовать росту новообразований и вызывать рак.
Присутствие высоких уровней цианотоксинов в рекреационных и питьевых водах может вызывать у человека широкий спектр симптомов (таблица 1), включая повышение температуры, головную боль, мышечные и суставные боли, кожные волдыри, желудочные спазмы, понос, рвоту, язвы в ротовой полости и аллергические реакции. Данные эффекты могут проявляться в течение нескольких минут или дней после воздействия. В некоторых случаях могут происходить приступы, печеночная недостаточность, остановка дыхания и (редко) смерть. Существуют доказательства, что долгосрочное воздействие микроцистинов и цилиндроспермопсина может способствовать росту новообразований и вызывать рак.
Существует много документированных сообщений о гибели собак, птиц и скота во всех регионах мира в результате потребления поверхностных вод с цветением цианобактерий. Нечасто также документирована гибель людей. Внутривенное введение воды, содержащей микроцистины, в центре почечного диализа в Бразилии в 1996 году привело к смерти 50 человек.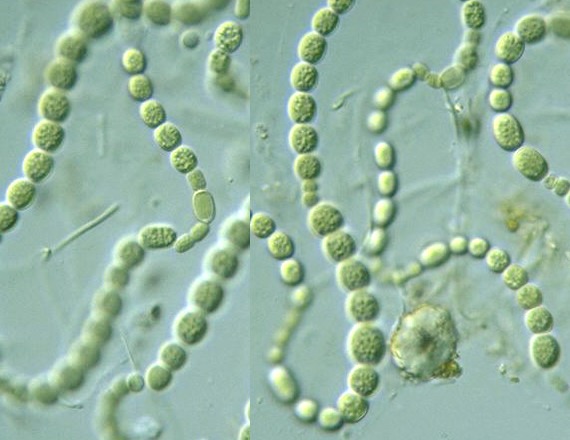 Действительный риск воздействия низких концентраций цианотоксинов в питьевой воде и долгосрочные эффекты воздействия этих токсинов точно неизвестны.
Действительный риск воздействия низких концентраций цианотоксинов в питьевой воде и долгосрочные эффекты воздействия этих токсинов точно неизвестны.
Методы проверки
Таблица 2 описывает методы, применимые для измерения концентрации цитотоксинов в воде.
Существуют коммерческие полевые наборы тестов для использования в качестве инструмента проверки присутствия или отсутствия специфических цианотоксинов в источниках водоснабжения. Проверочные наборы (ELISA1наборы) пригодны для определения микроцистинов, цилиндроспермопсина и сакситоксина, но в настоящее время непригодны для определения анатоксина-а. Хотя они предоставляют экспрессные результаты, в целом данные наборы имеют ограничения по точности, чувствительности и специфичности. В случае, если проверочный тест положительный, система водоснабжения должна направить пробы в лабораторию, способную количественно измерить концентрации специфичных цианотоксинов, используя более точные методики, такие как жидкостная хроматография, объединенная с масс-спектрометрией (LC/MS).
Очистка воды от цианотоксинов и управление цветением
При обнаружении в поверхностной воде, используемой для питьевой водоподготовки, цианобактерий и/или их токсинов операторы станции питьевой водоподготовки могут применить несколько способов для их удаления или инактивации. Некоторые способы водоподготовки эффективны в отношении определенных цианотоксинов, но не в отношении других групп цианотоксинов. Поэтому операторы станций питьевой водоподготовки должны знать типы роста и доминирующие во время цветения виды цианобактерий, свойства цианотоксинов (например, внутриклеточные или внеклеточные) и наиболее эффективные технологии водоподготовки. Применение ошибочного технологического процесса на специфическом этапе водоподготовки может повредить клетки и привести больше к высвобождению, чем удалению цианотоксинов.
В таблице 3 приведены данные об эффективности различных типов технологий водоподготовки в удалении неповрежденных клеток и процессах водоподготовки, которые эффективны в удалении внеклеточных растворенных цианотоксинов некоторых наиболее важных цианобактерий. Производители питьевой воды поощряются к мониторингу подготовленной воды, чтобы гарантировать удаление цианотоксинов.
Производители питьевой воды поощряются к мониторингу подготовленной воды, чтобы гарантировать удаление цианотоксинов.
Производители питьевой воды могут применять различные стратегии управления, связанные с цветением цианобактерий, чтобы избежать высвобождения цианотоксинов в воду. Например, те станции питьевой водоподготовки, которые имеют более одного источника водоснабжения, могут их менять. Другой альтернативой управления может быть изменение глубины водозабора, чтобы избежать попадания на станцию загрязненной воды и клеток цианобактерий.
Для уменьшения концентраций веществ, влияющих на привкус и запах воды, уменьшения количества дрейссен и прочих загрязнителей часто применяется предварительная обработка воды окислителем на входе процесса водоподготовки. Однако использование данного метода вызывает определенное беспокойство в отношении разрыва клеток и высвобождения токсинов. Обработка сульфатом меди и озоном на входе процесса водоподготовки удаляет водорослевое цветение, но она не рекомендуется из-за риска лизиса клеток водорослей. Хлорирование, в дополнение к разрушению клеток, может приводить к образованию в процессе очистки питьевой воды побочных продуктов дезинфекции. Если необходима предварительная обработка окислителем, то для перманганата калия (KMnO4) продемонстрирована эффективность в удалении клеток Micocystis без высвобождения токсина. В дополнение рекомендуется применять порошковый активированный уголь (PAC), чтобы удалить любые потенциально высвободившиеся токсины.
Хлорирование, в дополнение к разрушению клеток, может приводить к образованию в процессе очистки питьевой воды побочных продуктов дезинфекции. Если необходима предварительная обработка окислителем, то для перманганата калия (KMnO4) продемонстрирована эффективность в удалении клеток Micocystis без высвобождения токсина. В дополнение рекомендуется применять порошковый активированный уголь (PAC), чтобы удалить любые потенциально высвободившиеся токсины.
Показано, что стандартный процесс очистки питьевой воды (коагуляция, флоккуляция, седиментация и фильтрация) может быть эффективен в удалении внутриклеточных токсинов. Коагуляция, флоккуляция и флотация растворенным воздухом (DAF) более эффективны, чем седиментация. Микрофильтрация и ультрафильтрация в высшей степени эффективны в удалении неповрежденных клеток цианобактерий. Когда наблюдается цветение, и клетки цианобактерий проходят через фильтры, следует чаще проводить обратную промывку, чтобы снизить риск высвобождения токсинов в воду.
Для удаления внеклеточных токсинов производители питьевой воды могут использовать активированный уголь, мембранную фильтрацию и химическую инактивацию (ультрафиолетовое облучение, дезинфектанты и оксиданты). Порошковый активированный уголь (PAC) и гранулированный активированный уголь (GAC) эффективны при абсорбции микроцистина и цилиндроспермопсина, хотя разновидности микроцистина могут иметь разные эффективности абсорбции.
Эффективность работы активированного угля зависит от концентрации токсина, а также от дозы и происхождения активированного угля. Для оценки эффективности различных типов PCA рекомендуется проводить поверхностные тесты яса с применением угля с наибольшей способностью удаления целевых загрязнителей. GAC-фильтры эффективны в удалении микроцистинов, если они должным образом заменяются или регенерируются при высоком проскоке общего органического углерода. Обычно для эффективного удаления токсинов требуются повышенные концентрации активированного угля, для полного удаления токсинов может быть необходима повторная обработка. Нанофильтрация и обратный осмос эффективны в удалении цилинроспермопсина и микроцистина. Но в любом случае рекомендуется проводить пилотные исследования, поскольку эффективность удаления зависит от распределения размера пор мембраны и качества воды
Нанофильтрация и обратный осмос эффективны в удалении цилинроспермопсина и микроцистина. Но в любом случае рекомендуется проводить пилотные исследования, поскольку эффективность удаления зависит от распределения размера пор мембраны и качества воды
Обработка ультрафиолетовым излучением эффективна в разрушении клеток с микроцистином, анатоксином-а и цилиндроспермопсином. Однако в связи с тем, что для этого требуются более высокие дозы, чем применяемые на практике, данная обработка становится практически нереальной. УФ-обработка использовалась с катализатором (оксидом титана), чтобы разрушить токсины окислением. Но эффективность данного процесса в значительной степени зависела от содержания органики в воде. Оксиданты, такие как хлор, озон и перманганат калия, являются эффективными способами инактивации микроцистина, однако эффективность хлора зависит от рН. Различные цианоксины реагировали на хлор по-разному, например, доказано, что хлор неэффективен для инактивации анатоксина-а. Когда же рН ниже 8, хлор эффективен для инактивации микроцистина и цилиндроспермопсина. Озон — хороший оксидант для микроцистинов, анатоксина-а и цилиндроспермопсина, но его эффективность также зависит от рН, присутствие органического вещества может влиять на эффективность озонирования. Хотя окисление озоном микроцистина не зависит от рН, эффективность озона зависит от рН (от 7 до 10) в отношении анатоксина-а и цилиндроспермопсина (от 4 до 10). Перманганат калия эффективен в окислении микроцистина и анатоксина-а (при рН от 6 до 8), но не очень реакционноспособен в отношении цилиндроспермопсина. Обработка хлораминами и оксидом хлора неэффективна для микроцистина, анатоксина-а и цилиндроспермопсина.
Озон — хороший оксидант для микроцистинов, анатоксина-а и цилиндроспермопсина, но его эффективность также зависит от рН, присутствие органического вещества может влиять на эффективность озонирования. Хотя окисление озоном микроцистина не зависит от рН, эффективность озона зависит от рН (от 7 до 10) в отношении анатоксина-а и цилиндроспермопсина (от 4 до 10). Перманганат калия эффективен в окислении микроцистина и анатоксина-а (при рН от 6 до 8), но не очень реакционноспособен в отношении цилиндроспермопсина. Обработка хлораминами и оксидом хлора неэффективна для микроцистина, анатоксина-а и цилиндроспермопсина.
Образование побочных продуктов дезинфекции является другой вероятной проблемой при использовании озона, сульфата железа и хлора при высоких концентрациях бромида в воде. Тем не менее результаты исследований воздействия хлорирования на внутриклеточные токсины и результирующее образование побочных продуктов дезинфекции противоречивы. Большинство исследований полагают, что в течение цветения следует в идеале избегать предварительного хлорирования до тех пор, пока должные уровни контактного времени не будут гарантировать полного эффективного окисления разрушенных цианобактерий.
Разработка плана действий в чрезвычайных ситуациях
Организации, управляющие водными запасами, должны разработать план действий в чрезвычайных ситуациях при цветении цианобактерий. Большинство массовых развитий водорослей нетоксично, и план должен предусматривать, как определить риск, связанный с каждым событием. Элементы такого плана должны определять, где и когда отбираются пробы; частоту отбора проб; объем проб; когда отбираются пробы на наличие клеток цианобактерий или специфических токсинов или одновременно на наличие клеток и токсинов; какой аналитический тест используется для наблюдения за ситуацией; условия, при которых необходимо отослать в выбранную лабораторию пробу (пробы) для подтверждения. Производители питьевой воды должны также разработать план альтернативных действий до получения результатов аналитического контроля, включая способ (способы) очистки для применения при положительных результатах анализов, а также определение процесса общественного оповещения.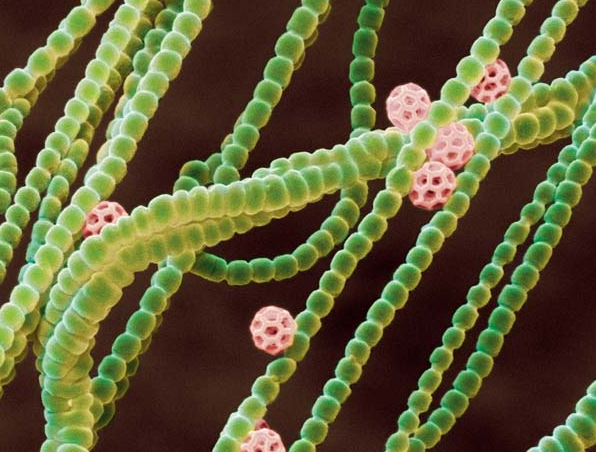 Для разработки таких планов может использоваться глава 6 («Оценка ситуации, планирование и управление») из издания ВОЗ «Токсичные цианобактерии в воде: руководство по последствиям для общественного здоровья, мониторингу и управлению».
Для разработки таких планов может использоваться глава 6 («Оценка ситуации, планирование и управление») из издания ВОЗ «Токсичные цианобактерии в воде: руководство по последствиям для общественного здоровья, мониторингу и управлению».
Перевод — Владимир Ларин, кандидат биологических наук, начальник отдела биологических методов анализа ЗАО «РОСА»
Фото: ufaleyadmin.ru
Журнал «Вода Magazine» №4 (92), 2015 г.
Cyanobacteria — Bilder und stockfotos
12.623Bilder
- Bilder
- Fotos
- Grafiken
- Vektoren
- Videos
DUCHSTEREN
. Odersuchen Sie nach blaualge, um noch mehr faszinierende Stock-Bilder zu entdecken.
nostoc sp. фото и изображения цианобактерий под микроскопом
Nostoc SP. Альген под микроскопом Sicht
Альген под микроскопом Sicht
Nostoc ist eine Gattung von Cyanobakterien, die in verschiedenen Umgebungen vorkommen und Kolonien bilden, die aus Filamenten von moniliformen Zellen in einer gallertartigen Hülle bestehen.
носток сп. algen unter mikroskopischer betrachtung, cyanobakterien, giftiges wasser — cyanobacteria stock-fotos und bilder
Nostoc sp. Algen unter mikroskopischer Betrachtung,…
нитчатые цианобактерии, oscillatoria arten, aufnahme — cyanobacteria stock-fotos und bilder
нитчатые цианобактерии, Oscillatoria Arten, Aufnahme
Mikroaufnahme von fadenförmigen Cyanobakterien oder Blaualgen,
spirulina trockenttabletten. таблетки спирулины в tassen auf grünem hintergrund. вегетарианские омега дрей. pflanzliche quelle von aminosäuren und omega 3.omega drei альтернативный quelle. супер эссен. — цианобактерии фото и изображения
Spirulina Trockenttabletten. Spirulina Pillen in Tassen auf grünem
die 3d Colonie von Bacterien in der umwelt — цианобактерии фото и изображения
Die 3d Colonie von Bacterien in der Umwelt
Бактериальные и вирусные инфекции. 3D-Render-Illustrationskonzept
3D-Render-Illustrationskonzept
cyanobacteria aufnahme — cyanobacteria stock-fotos und bilder
cyanobacteria Aufnahme
Cyanobakterien oder Blaualgen,
makroskopische energie solar-zu-kraftstoff grün cyanobacterium zelle banner hintergrund — cyanobacteria stock-fotos und bilder
Makroskopische Energie Solar-zu -Kraftstoff grün Cyanobacterium…
Mikrobielle Biobrennstoff-Energiezelle in makroskopischer Ansicht. Концепт 3D-иллюстрацииМодель. Solar-to-Fuel-Algen Ethanol-Power und entwickeltes Cyanobakterium, Algenbakterien als zukünftige Zero-Waste-erneuerbare Energie
algenforschung В лаборатории, биотехнологии Wissenschaftskonzept — Cyanobacteria Stock -fotos und Bilder
Algenforschung в Laboratorien, Biotechnologches …
Algen -Algen -Algen -Deslerge Deslen -Deslen -Deslen -Deslen -Deslen -Deslerge Deslerge Deslerge Deslerge Deslerge Deslerge Deslerge Deslerge Deslerge Desleren -Desler. дер Wasseroberfläche. Blütenwasser als Hintergrund или Textur
дер Wasseroberfläche. Blütenwasser als Hintergrund или Textur
green wasser — cyanobacteria stock-fotos und bilder
Green Wasser
die küste auf der oberfläche des flusses ist mit einem pellel von blaugrünen algen beeckt, kopie raum. — цианобактерии фото и изображения
Die Küste auf der Oberfläche des Flusses ist mit einem Pellel…
cyanobakterien, auch bekannt als cyanophyta auf der wasseroberfläche. kann neurotoxine ein anderes toxin produzieren. algenblüten, die zu einer gefahr für mensch und tier werden können. — cyanobacteria stock-fotos und bilder
Cyanobakterien, auch bekannt als Cyanophyta auf der Wasseroberfläc
wissenschaftler entwickeln algenforschung. bioenergie, biokraftstoff, energieforschung — cyanobacteria стоковые фотографии и изображения
Wissenschaftler entwickeln Algenforschung. Bioenergie,…
menschliche hand und wasser mit einer schicht blaugrüner algen auf der wasseroberfläche. — cyanobacteria Stock-fotos und Bilder
Menschliche Hand und Wasser mit Einer Schicht blaugrüner Algen. ..
..
Питайя Smoothie in schüssel mit kiwi, banane, heidelbeeren und superfoods auf der oberseite — cyanobacteria Stock-fotos und Bilder
Pitaya Smoothie in Schüssel mit Kiwi , Banane, Heidelbeeren und…
wasserverschmutzung durch blühende blaualgen — цианобактерии, не связанные с проблемой мировоззрения. gewässer, flüsse und see mit schädlichen algenblüten. ökologiekonzept der verschmutzten natur. — цианобактерии фото и изображения
Wasserverschmutzung durch blühende Blaualgen — Cyanobakterien…
Wasserverschmutzung durch blühende Blaualgen — Cyanobakterien sind ein weltweites Umweltproblem. Gewässer, Flüsse und Seen mit schädlichen Algenblüten. Ökologiekonzept der verschmutzten Natur
photobioreactor — cyanobacteria stock-fotos und bilder
Photobioreactor
wirbelnde grün- und blaualgen (cyanobakterien) auf einem see, der mit dickem grünem wasser gefüllt ist, umgeben von bäumen an einem klaren blauen himmel — cyanobacteria stock-fotos und картинка
Wirbelnde Grün- und Blaualgen (Cyanobakterien) auf einem См. ,…
,…
nostoc cyanobacteria aufnahme — cyanobacteria stock-fotos und bilder
Nostoc cyanobacteria Aufnahme
vorsicht vor cyanobakterien in veldhuizen, de meerto9icht 0 bildertocyanobacteria stockn — cyanobacteria stock-fotos und bilder Cyanobacterien in Veldhuizen, De Meern
spirulina algenpulver in der Schüssel, альтернативная медицина для энтгифтунга, антиоксидантные lebensmittelzutaten — cyanobacteria stock-fotos und bilder
Spirulina Algenpulver in der Schüssel, Alternativmedizin zur…
spirulina algenpulver in der Schüssel, Alternativmedizin zur… cyanobacteria stock-fotos und bilder
Heidelbeer-Spirulina-Smoothie in Schüssel mit Physalis, Beeren…
спирулина в таблетках собрана в комбинированной геометрической форме. зеленая спирулина в таблетках на зеленом фоне в минималистическом стиле. — цианобактерии фото и изображения
Spirulina Pillen Muster auf grünem kombiniertem geometrischem…
Spirulina algenpulver in der Schüssel, alternativmedizin zur entgiftung, antioxidative lebensmittelzutaten — cyanobacteria stock-fotos und bilder
Spirulina Algenpulver in der Schüssel, grandprisemedizin 00002 Бассейн, Йеллоустонский национальный парк, Вайоминг. zeigt die heiße quelle und das wasser mit den bunten bakterien, die eine orange bis braune farbe geben. abfluss aus der heißen quelle mit den bunten bakterien. — цианобактерии фото и изображения
zeigt die heiße quelle und das wasser mit den bunten bakterien, die eine orange bis braune farbe geben. abfluss aus der heißen quelle mit den bunten bakterien. — цианобактерии фото и изображения
Гранд-призматический источник, бассейн Мидуэй-Гейзер, Йеллоустонский национальный…
блестящие блестки на зеленых водоемах — цианобактерии фото и изображения
блестящие пузыри на красных водоемах
судак (sander lucioperca) под водой. fleischfressende fische mit markierten flossen. унтер вассер гефанген. dunklehintergrund, steinige unterseite. — cyanobacteria stock-fotos und bilder
Судак (Sander lucioperca) унтер-Вассер. Fleischfressende Fische…
спирулина algenttabletten в прозрачной упаковке в einer reihe auf hellgrünem hintergrund. зеленая спирулина в таблетках. витамины и nahrungsergänzungsmittel. супер эссен. nahrungsergänzungsmittel — стоковые фотографии и изображения цианобактерий
Spirulina Algenttentabletten in Transparent Tassen in einer Reihe…
grüne spirulina smoothie schüssel — cyanobacteria stock-fotos und bilder
Grüne Spirulina Smoothie Schüssel
Green Spirulina Smoothie Bowl mit verschiedenen Toppings
спирулина в таблетках grüne grunded auf grund.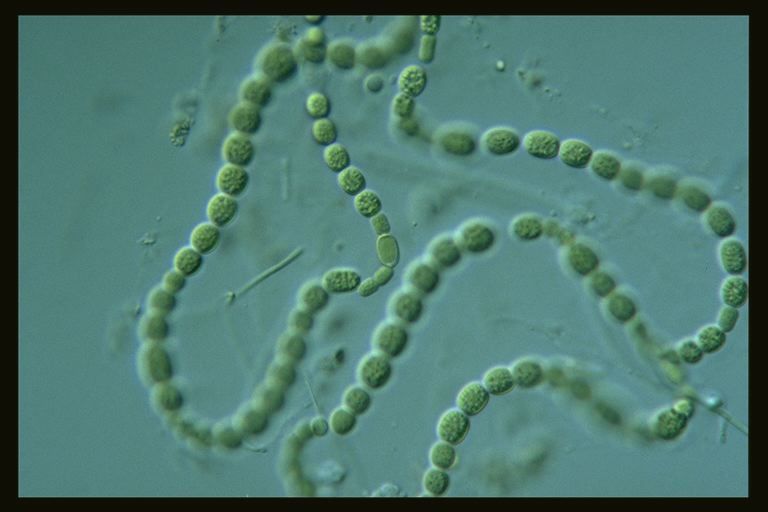 спирулина алгентная таблетка. витамины и nahrungsergänzungsmittel. супер эссен. nahrungsergänzungsmittel für einen gesunden lebensstil — cyanobacteria stock-fotos und bilder
спирулина алгентная таблетка. витамины и nahrungsergänzungsmittel. супер эссен. nahrungsergänzungsmittel für einen gesunden lebensstil — cyanobacteria stock-fotos und bilder
Spirulina Tabletten auf grünem Hintergrund. Спирулина…
спирулина-алген. хлорелла algen grüne tableten isoliert auf weißemhintergrund.algen nahrungsergänzungsmittel. таблетка порошка хлореллы. Альтернативная медицина и гомеопатия. суперэссенция — цианобактерии фото и фотография
Spirulina-Algen. Chlorella Algen grüne Tabletten isoliert auf weiß
weizengraspulver isoliert auf weißem hintergrund — cyanobacteria stock-fotos und bilder
Weizengraspulver isoliert auf weißem Hintergrund
denkmal für maximilian ii. в Мюнхене. — цианобактерии фото и изображения
Денкмаль для Максимилиана II. в Мюнхене.
Kleiner Fluss voller grünalgen — cyanobacteria stock-fotos und bilder
Kleiner Fluss voller Grünalgen
Kleiner Fluss voller Grünalgen and Heißen Sommertagen
колония зеленых зеленых альгенов в культуральной среде, микробиологическая среда.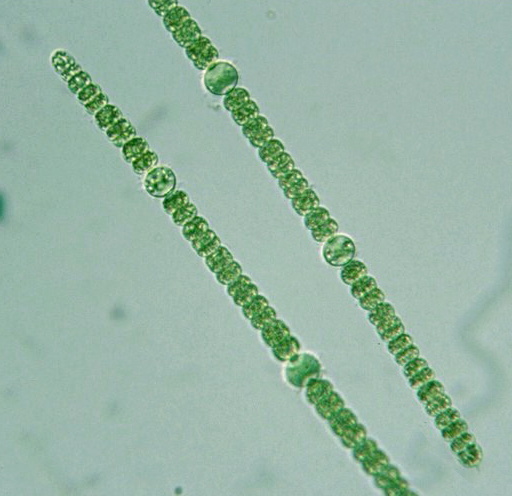 — cyanobacteria stock-fotos und bilder
— cyanobacteria stock-fotos und bilder
Colonie der blue grünen Algen in Kultur Medium Platte,…
cyanobacterien vektor-illustration. beschriftete bakterien interne struktur schema — cyanobacteria stock-grafiken, -clipart, -cartoons und -symbole
Цианобактерии Вектор-Иллюстрация. Beschriftete Bakterien…
Cyanobakterien-Vektor-Illustration. Gekennzeichnetes internes Strukturschema für Bildungsbakterien. Biologisches Blaualgendiagramm mit Carboxysom-, Thylakoid- und Phycobilisomen-Teilen innerhalb der Zelle.
blaugrüne algen — цианобактерии стоковые фото и фотографии
прудовой микроорганизм-автотроф — цианобактерии сток-график, -клипарт, -мультфильмы и -символ
Pond micro Organismen-Autotroph
strand in einem süßwassersee mit einer dichten blüte der grünalgen. — cyanobacteria stock-fotos und bilder
Strand in einem Süßwassersee mit einer dichten Blüte der Grünalgen
denkmal für maximilian ii. (maxmonument) в Мюнхене, Германия.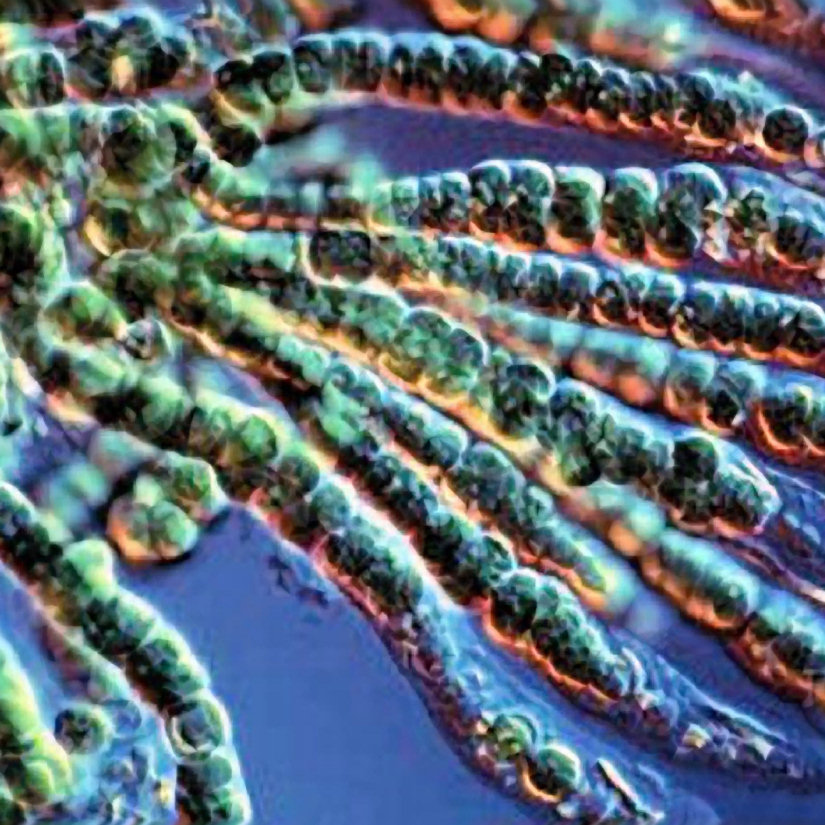 das denkmal zu ehren des bayerischen königs maximilian ii. wurde 1875 eingeweiht. — цианобактерии фото и фотографии
das denkmal zu ehren des bayerischen königs maximilian ii. wurde 1875 eingeweiht. — цианобактерии фото и фотографии
Denkmal für Maximilian II. (Maxmonument) в Мюнхене, Германия.
bunte gelbe flechten auf der alten steinmauer outdoor naturunschärfehintergrund — cyanobacteria stock-fotos und bilder
bunte gelbe Flechten auf der alten Steinmauer Outdoor Naturunschär
Xanthoria parietina auch gelbe Flechten genannt, die auf einer von der Herbstsonne beleuchteten Steinmauer wachsen
asiatische chinesische mittel erwachsene kurzhaarfrau, die ihr morgendliches aufwachen mit frischem obst-smoothie auf dem balkon genießt — cyanobacteria Stock-fotos und bilder
asiatische chinesische Mittel erwachsene Kurzhaarfrau, die ihr…
microalgen und cyanobakterien — cyanobacteria стоковые графики, -клипарты, -мультфильмы и -символы
Mikroalgen und Cyanobakterien
algenblüte im wasser des dnjepr in der ukraine. grüne farbe des wassers, verursacht durch eine schnelle zunahme oder ansammlung von algen.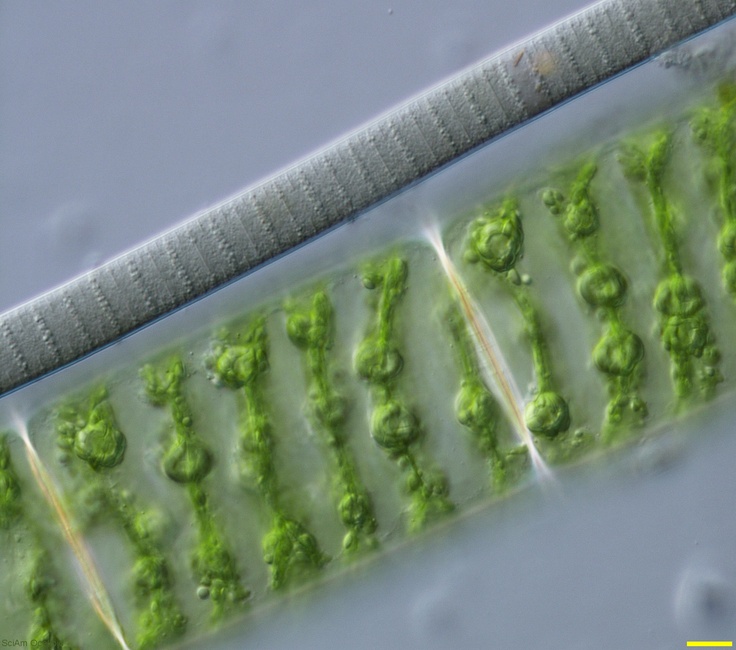 экологическая катастрофа. wasserverschmutzung. natürlicher grüner abstrakter intergrund — cyanobacteria stock-fotos und bilder
экологическая катастрофа. wasserverschmutzung. natürlicher grüner abstrakter intergrund — cyanobacteria stock-fotos und bilder
Algenblüte im Wasser des Dnjepr in der Ukraine. Grüne Farbe des…
Spirulina Smoothie isoliert auf Weißem Hintergrund — cyanobacteria stock-fotos und bilder
Spirulina Smoothie isoliert auf weißem Hintergrund
Hochwinkelansicht eines mit Spirulina-Smoothie gefüllten Trinkglases und einer Schüssel mit Spirulina-Pulver, isoliert auf weißem Hintergrund. Цифровая фотокамера Hochauflösende 42Mp с Sony A7rII и Sony FE 90mm f2.8 Makro G OSS Objektiv
blau – grüne alge – cyanobacteria stock-fotos und bilder
blau – grüne Alge
Blaualge infiziert smoothiebacteria fluss
ges фото и фотографии
Синяя чаша Spirulina Smoothie Bowl
Голубая чаша для смузи со спирулиной и шванцем Marmaid
голубых и зеленых желтков, которые были заморожены.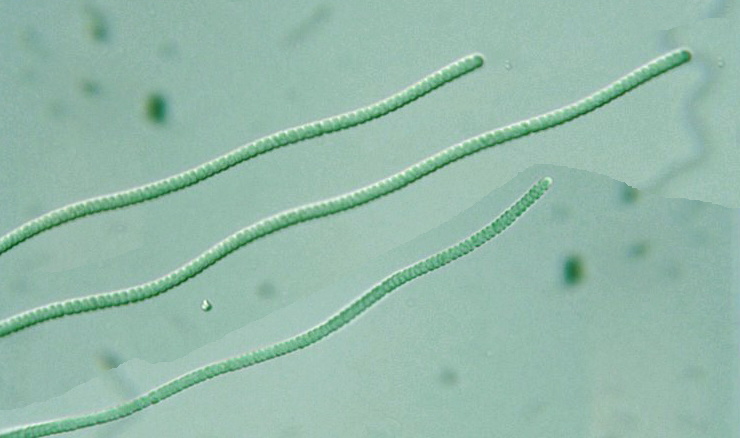 заражение дурным алгеном. umweltverschmutzung. экологическая катастрофа. — cyanobacteria stock-fotos und bilder
заражение дурным алгеном. umweltverschmutzung. экологическая катастрофа. — cyanobacteria stock-fotos und bilder
Schmutziges blaues und grünes Wasser im Stausee. Контаминация…
Цветение цианобактерий Фотогалерея | Департамент природных ресурсов Миссури
Цветы могут быть разных цветов, например ярко-зеленого, сине-зеленого, белого, красного или желтовато-коричневого. Вода может выглядеть как гороховый суп, или поверхность может выглядеть так, как будто на воду пролили краску. Они могут иметь или не иметь густую пену или пену. Многие цианобактерии могут тонуть или плавать, чтобы контролировать свое положение в толще воды, поэтому может казаться, что цветение движется и быстро меняется. Другие факторы, такие как ветер, дождь, пробуждение от развлекательных мероприятий и течения, также могут повлиять на место цветения или на то, как оно появляется.
Так как же узнать, цианобактерии у вас или нет? Вы всегда можете связаться со специалистом, заполнив Форму уведомления о подозрении на вредоносное цветение водорослей (HAB) MO 780-2827 . Член группы реагирования будет рад помочь вам определить, стоит ли вам беспокоиться по поводу цветения. Вы также можете просмотреть фотогалерею ниже, чтобы увидеть примеры типичного цветения Миссури, или перейти по ссылкам ниже, чтобы увидеть дополнительные примеры цианобактерий и водорослей, отличных от сине-зеленых.
Член группы реагирования будет рад помочь вам определить, стоит ли вам беспокоиться по поводу цветения. Вы также можете просмотреть фотогалерею ниже, чтобы увидеть примеры типичного цветения Миссури, или перейти по ссылкам ниже, чтобы увидеть дополнительные примеры цианобактерий и водорослей, отличных от сине-зеленых.
- Полевой и лабораторный справочник по пресноводным цианобактериям, вредным цветениям водорослей для общин коренных американцев и коренных жителей Аляски
- Фото не сине-зеленых водорослей
 jpg» data-sub-html=»»>
jpg» data-sub-html=»»> jpg» data-sub-html=»»>
jpg» data-sub-html=»»> jpg» data-sub-html=»»>
jpg» data-sub-html=»»> jpg» data-sub-html=»»>
jpg» data-sub-html=»»> jpg» data-sub-html=»»>
jpg» data-sub-html=»»>Чтобы просмотреть всю галерею, нажмите на любое изображение.
Видео | |||
Большой побег | |||
Пастбище на циано | |||
Спирулина в движении | |||
Прядение Спирулина | |||
Изображения | |||
Anabaena affinis , 640X | |||
Anabaena circinalis — Измерение невозможно. | |||
Anabaena doliolum | |||
Anabaena flos-aquae увеличенный 2500X | |||
Anabaena «laxa» — Идентификатор этого комплекса, конечно, незначителен — см. следующую запись. Измерение недоступно. вероятно 400X | |||
Anabaena «laxa» — Из того же образца, что и выше, несколько недель спустя. Обратите внимание, что акинет теперь примыкает к гетероцисте. Теперь это ключи к Анабаена « цилиндрическая «. 400Х | |||
Anabaena scheremetievi — фазовый контраст 400X | |||
Anabaena scheremetievi — Виден легкий намек на ножны. 400Х | |||
Anabaena scheremetievi — 400X | |||
Anabaena sphaerica , 1600X | |||
Anabaena sphaerica — 400X | |||
Anabaena sphaerica | |||
Anabaena spiroides — Линия 20 мкм. | |||
Anabaena torulosa — 400X | |||
Anabaena torulosa — Синекокок? Нет, это Anabaena torulosa akinetes. 400Х | |||
Anabaena variabilis | |||
Anabaena sp. — (Неопознанный) Никаких предположений о виде. 400X | |||
Anabaena sp. — (Неопознанный) Никаких предположений о виде. 400Х | |||
Anabaena sp. — (Неопознанный) Никаких предположений о виде. 400Х | |||
Anabaena sp. — (Необычные и искривленные) Акинеты круглые, так что, вероятно, это не A. circinalis . Может быть A. spiroides в необычном состоянии. 200X. | |||
Anabaena sp. | |||
Anabaena sp. — Вероятно A. scheremievi , но акинеты отсутствовали. Намек на ножны едва заметен. 400Х | |||
Anabaena sp. — Вероятно A. circinalis , но акинеты отсутствовали. 1000X | |||
Anabaena sp. — Возможно A. viguieri , но акинеты слишком широкие. 400X. | |||
Anabaena sp., из пресноводного озера в Орегоне, 100X | |||
Anabaena sp., из пресноводного озера в Орегоне, 100X | |||
Anabaena sp., из пресноводного озера в Орегоне, 200X | |||
Anabaena sp., из пресноводного озера в Орегоне, 100X. | |||
Anabaena sp. | |||
Anabaena sp. | |||
Anabaena sp. | |||
Anabaena sp. | |||
Anabaena sp. с бактериями-симбионтами (возможно Zoogloea ) вокруг гетероцист | |||
Anabaenopsis circleis — Хорошо видна центральная пара гетероцист. 200Х | |||
Anabaenopsis circleis — В центре трихомы видна развивающаяся пара гетероцист. Обратите внимание на прикрепленные к нему бактерии. 400X (фазовый контраст) | |||
Анабаенопсис круговой — То же, что и выше, но без фазового контраста. | |||
Anabaenopsis sp. цветение на озере Бедетти, Санто-Томе, Санта-Фе, Аргентина | |||
Anabaenopsis sp. цветение на озере Бедетти, Санто-Томе, Санта-Фе, Аргентина | |||
Anabaenopsis sp. цветение на озере Бедетти, Санто-Томе, Санта-Фе, Аргентина | |||
Anacystis nidulans ПЭМ полутонких срезов (200 нм). См. протокол. | |||
Anacystis nidulans ПЭМ полутонких срезов (200 нм) См. протокол. | |||
Anacystis nidulans ПЭМ полутонких срезов (300 нм) См. протокол. | |||
Anacystis nidulans ПЭМ полутонких срезов (300 нм) См. протокол. | |||
Anacystis nidulans ПЭМ полутонких срезов (500 нм) См. | |||
Anacystis nidulans ПЭМ полутонких срезов (500 нм) См. протокол. | |||
Aphanizomenon flos-aquae — Хорошо видны акинеты и гетероцисты. Измерение недоступно. вероятно 400X | |||
Aphanizomenon flos-aquae — Колония. Измерение недоступно. вероятно 100X | |||
Aphanizomenon flos-aquae — Колония. Акинеты отсутствуют. 400Х | |||
Aphanizomenon flos-aquae штамм 1401/5. Автофлуоресцентное изображение. | |||
Aphanizomenon flos-aquae штамм 1401/5. Автофлуоресцентное изображение. | |||
Aphanizomenon flos-aquae штамм 1401/5. Автофлуоресцентное изображение. | |||
Артронема африканская | |||
Arthrospira jenneri — Поперечные стенки закрыты цитоплазматическими гранулами (извините). | |||
Beggiatoa alba — 400X | |||
Beggiatoa alba — Обрезанная часть изображения выше с более высоким разрешением. | |||
Цветение в водах плотины Хартебееспоорт в Южной Африке | |||
Цветение Microcystis aeruginosa и Anabaena circinalis на реке Сент-Джонс, Флорида | |||
Цветение Aphanizomenon flos aquae в озере в Суррее, Великобритания | |||
Блум | |||
Цветок Microcystis в Кресент-Лейк, Флорида | |||
Блум в Докторском озере | |||
Цветение в озере Поншартрейн, штат Луизиана, олигогалинном эстуарии | |||
Цветок Microcystis aeruginosa и Anabaena circinalis загрязнено лодкой на реке Сент-Джонс, Флорида | |||
Цветение в сельскохозяйственных условиях в штате Миссури | |||
Цветение в сельскохозяйственных условиях в штате Миссури | |||
Цветение в сельскохозяйственных условиях в штате Миссури | |||
Цветение в сельскохозяйственных условиях в штате Миссури | |||
Цветение в сельскохозяйственных условиях в штате Миссури | |||
Anabaena и Aphanizomenon — разлагающаяся пена в шотландском пресноводном озере | |||
Anabaena и Microcystis — цветение в шотландском пресноводном озере | |||
Borzia trilocularis | |||
Borzia sp. | |||
Калотрикс Брауни | |||
Calothrix sp. ПКК 7103 | |||
Calothrix sp. | |||
Calothrix sp. показаны терминальные гетероцисты | |||
Calothrix sp. | |||
Calothrix sp. | |||
Calothrix sp. | |||
Калотрикс зр. | |||
Chloroflexus sp. — Неизвестный фитопланктон из рыбного пруда. Предположение: Chloroflexus . 400Х | |||
Chloroflexus sp. — Более широкое изображение с более высоким разрешением вышеприведенного образца. | |||
Chroococcidiopsis kashayi | |||
Chroococcidiopsis kashayi | |||
Chroococcidiopsis sp. | |||
Chroococcus lithophilus | |||
Chroococcus sp. | |||
Сбор цианобактерий с большого цветка | |||
Cryptozoon proliferum — из Квебека | |||
Cryptozoon proliferum — из Квебека | |||
Цианобактериальный налет | |||
Цианобактерии кедорум | |||
Cyanobacterium notatum | |||
Cyanosarcina sp. | |||
Cylindrospermopsis raciborskii — Видны гетероцисты и акинеты. 400Х | |||
Очень старая лабораторная культура Cyanothece sp. образует прочную крышу для удержания влаги. | |||
Цианотеки sp. — электронная микрофотография, показывающая углеводные гранулы | |||
Cylindrospermopsis raciborskii — фазовый контраст 400X | |||
Цилиндроспермопсис расиборский — Видна гетероциста. 400Х | |||
Cylindrospermopsis raciborskii — Akinetes (только) видимые. 400-кратный фазовый контраст | |||
Cylindrospermopsis raciborskii — Видна гетероциста. | |||
Cylindrospermum muscicola — 400X | |||
Cylindrospermum muscicola — Сюрприз! Это не Pseudanabaena , а C. muscicola , без акинет, но с развивающейся гетероцистой. 400-кратный фазовый контраст | |||
Cylindrospermum sp. ул. ПКК 7417 | |||
Cylindrospermum sp. | |||
Cylindrospermum sp., 1600X | |||
Cylindrospermum sp., 1600X | |||
Eubacterium — Неизвестная бактерия наносит визит Euglena . 1000X | |||
Fischerella sp. ул. ПКК 7414 | |||
Fischerella sp. | |||
Geitlerinema sp. из системы переработки целлюлозно-бумажных отходов в Айдахо | |||
Girvanella sp. в онколитах водорослей из нижнекембрийского известняка Мул Спринг, Ваукоба Спринг, Национальный парк Долины Смерти | |||
Gloeobacter sp. ул. ПКК 8105 | |||
Gloeobacter sp. | |||
Gloeocapsa sp. вместе с двумя водорослями | |||
« Gloeotrichia » — хорошо видны акинеты и гетероцисты. Измерение недоступно. 400X | |||
« Глоотрихия » — Колония. Измерение недоступно. 100Х | |||
Gloeotrichia echinulata — колонии размером с булавочную головку в шотландском пресноводном озере | |||
Ягайнема пренебрежительная | |||
Комвофорон сп. | |||
Leptolyngbya africana | |||
Leptolyngbya crispata | |||
Leptolyngbya foveolarum f maio | |||
Leptolyngbya foveolarum | |||
Leptolyngbya foveolarum | |||
Leptolyngbya fragilis | |||
Leptolyngbya fragilis | |||
Leptolyngbya fragilis | |||
Лептолингбья Фригида | |||
Лептолингбья Шмидлей | |||
Лептолингбья Скотти | |||
Leptolyngbya sp. | |||
Lyngbya aestuarii | |||
Lyngbya majuscula | |||
Lyngbya sp. | |||
Lyngbya sp. демонстрирует классическую оболочку и терминальную дифференцировку в нижней нити | |||
Lyngbya sp. с изображением спирали на конце шпильки | |||
Lyngbya sp. — (Неопознанный) Никаких предположений о виде. 200Х | |||
Lyngbya sp. — (Неопознанный) Никаких предположений о виде. Обратите внимание, что у одного нет видимой оболочки. 200Х | |||
Lyngbya sp. — Тот же образец, что и выше. 400-кратный фазовый контраст | |||
Lyngbya sp. | |||
Lyngbya sp. — Общественное скопление вышеуказанных видов. 200Х | |||
Lyngbya sp. | |||
Lyngbya sp. | |||
Lyngbya sp., две клетки покидают скорлупу | |||
Lyngbya sp. | |||
Lyngbya sp. в 100 х. | |||
Сообщество ковриков Вязаные нити цианобактерий из зеленого слоя микробного коврика из Грейт-Сиппевиссет Солтмарш, Фалмут, Массачусетс | |||
Mat Community Поперечное сечение микробного мата из Great Sippewisset Saltmarsh, Falmouth, MA | |||
Merismopedia — Колония Merismopedia ( Agmenellum [Drouet], Synechocystis кластер Low GC [Waterbury and Rippka]). | |||
Merismopedia elegans , 640X | |||
Microcoleus cataractarum | |||
Microcoleus chthonoplastes | |||
Microcoleus chthonoplastes | |||
Microcoleus sociatus | |||
Microcoleus steenstrupii | |||
Microcoleus subtorulosus | |||
Microcoleus vaginatus | |||
Microcoleus vaginatus | |||
Microcoleus sp. | |||
Microcoleus sp. — Один из аномальных Microcoleus / Symploca / Lyngbya / Угадай . | |||
Микрокорка на почве | |||
Microcystis aeruginosa увеличено 2500X | |||
Microcystis aeruginosa штамм PCC 7806. Автофлуоресцентное изображение. | |||
Microcystis aeruginosa штамм PCC 7806. Автофлуоресцентное изображение. | |||
Microcystis aeruginosa штамм PCC 7806. Автофлуоресцентное изображение. | |||
Microcystis aeruginosa штамм PCC 7806. Автофлуоресцентное изображение. | |||
Microcystis aeruginosa штамм PCC 7806. Автофлуоресцентное изображение. | |||
Microcystis aerogenosa штамм 1450/10 в очень старых (6 месяцев) культурах в стационарной фазе, визуализированных с помощью автофлуоресценции. | |||
Microcystis aerogenosa штамм 7806 в очень старых культурах в стационарной фазе, визуализированных с помощью автофлуоресценции. Обратите внимание на временную особенность «яркого удара». 1000X | |||
Microcystis aerogenosa штамм 7806 в очень старых (100 дней) культурах с неподвижной фазой, визуализированных с фазовым контрастом. Обратите внимание на многоплоскостное деление. 1000X | |||
Microcystis aerogenosa штамм 7806 в очень старых (100 дней) культурах с неподвижной фазой, визуализированных с фазовым контрастом. Обратите внимание на узкую оболочку. 1000X | |||
Микроцистис аэрогеноза . 100Х | |||
Microcystis sp. цветение | |||
Microcystis sp. | |||
Microcystis sp. цветение с высоты птичьего полета | |||
Цветение Microcystis aeruginosa , озеро Мокоан, Виктория, Австралия. | |||
Microcystis sp. | |||
Microcystis sp. — Отдельные колонии. 200X | |||
Microcystis sp., цветение, Grandview Garden Park, Пекин | |||
Смесь — Oscillatoria cf. chalybea , Planktothrix agardhii и Anabaena spiroides . 200Х | |||
Смесь — Oscillatoria cf. chalybea , Planktothrix agardhii и Arthrospira jenneri . 200Х | |||
Смесь, включающая Thiothrix и Oscillatoria sp. | |||
Смесь, включающая Spirulina и Microcoleus sp. и диатомовые вместе с Thiopedia sp. | |||
Смесь цианобактерий из сообщества микробных матов | |||
Таинственный жук — действительно странный. Spirulina major внутри пачки. 400Х | |||
Mystery Bug — Неизвестная нить. Он сделан из диатомовых водорослей, десмидов, цианобактерий или неизвестного? 400Х | |||
Загадочный жук A — Planktothrix agardhii с набухшей терминальной клеткой. 200X. Для Mystery Bugs от A до E было высказано предположение, что набухшие терминальные клетки представляют собой плодовые тела (спорангии) грибов (хитридов), поскольку наблюдались гифы, идущие по длине подобных трихом. | |||
Загадочный жук B — Planktothrix agardhii с двойной вздутой терминальной клеткой. | |||
Загадочный жук C — Еще один странный Planktothrix agardhii . 400-кратный фазовый контраст. См. Mystery Bug A. | |||
Загадочный жук D — еще один странный Planktothrix agardhii . 400-кратный фазовый контраст. См. Mystery Bug A. | |||
Mystery Bug E — То же, что и выше, но без фазового контраста. 400X. См. Mystery Bug A. | |||
Загадочные волнистые цианобактерии | |||
Загадочные цианобактерии в колонии | |||
Загадочные цианобактерии с везикулами | |||
Тайна 13 | |||
Тайна 14 | |||
Тайна 15 | |||
Тайна 16 | |||
Nodularia spumigena увеличенный 1250X | |||
Nodularia sp. | |||
Nostoc (Anabaena) azollae — Хорошо видна гетероциста. 400Х | |||
Nostoc (Anabaena) azollae — Видны многочисленные акинеты. 400Х | |||
Коммуна Носток | |||
Носток линкия | |||
Nostoc muscorum | |||
Nostoc punctiforme | |||
Nostoc sp. — Возможен росток. 400X | |||
Nostoc sp. — Возможная колония или это странный тип Anabaena spherica ?. Единственное, в чем можно быть уверенным, так это в том, что ни в чем нет уверенности. 200Х | |||
Nostoc sp. | |||
Nostoc sp. при увеличении 10X | |||
Nostoc sp. | |||
Nostoc sp. | |||
Nostoc sp. | |||
Nostoc sp. | |||
Nostoc sp. | |||
Nostochopsis sp., клетки и разветвления | |||
Nostohopsis sp., общая структура | |||
Nostohopsis sp., детальное изображение | |||
Nostohopsis sp., гетероциста на ножке | |||
Nostochopsis sp., интеркалярная гетероциста | |||
Oscillatoria agardhii — Автофлуоресцентное изображение. | |||
Oscillatoria ср. chalybea — С Planktothrix agardhii и, возможно, Raphidiopsis sp. 200Х | |||
Oscillatoria « chlorina » — кавычки отражают недовольство автора идентификацией цианобактерий по цвету. 400Х | |||
Oscillatoria geminata — 400X | |||
Oscillatoria limnetica | |||
Oscillatoria limosa , 640X | |||
Oscillatoria margaritifera | |||
Осциллятория принцепс — 200X | |||
Осциллятория принцепс — 100X | |||
Oscillatoria Princeps — Встреча с друзьями. | |||
Oscillatoria splendida — фазовый контраст 400X | |||
Oscillatoria subuliformis | |||
Oscillatoria sp. — Знаменитый «большой Oscillatoria «. Виды не известны. Это может быть неописано. 200Х | |||
Oscillatoria sp. — Обратите внимание на гранулы в терминальной клетке вместе с ее утолщенной мембраной. 400Х | |||
Oscillatoria sp. — При ярком свете его поразительные гранулы становятся более заметными. Обратите внимание на спиральное расположение гранул. 100Х | |||
Oscillatoria sp. — Тот же образец, что и выше. 200X | |||
Oscillatoria sp. — При малом увеличении. 40Х | |||
Oscillatoria sp. | |||
Oscillatoria sp. | |||
Oscillatoria sp., 640X | |||
Oscillatoria sp., вероятно, гормогониум, из системы переработки целлюлозно-бумажных отходов в Бразилии | |||
Otlypothrix brevis | |||
Phormidium aerogineo-coerula | |||
Phormidium diguetti | |||
Phormidium hennigsii | |||
Phormidium jenkelianum | |||
Phormidium jenkelianum | |||
Формидиум ливидум | |||
Формидиум молле | |||
Phormidium pachydermaticum | |||
Phormidium pristleyi | |||
Phormidium retzii — Предварительная идентификация. | |||
Phormidium uncinatum — Предварительная идентификация. 400-кратный фазовый контраст | |||
Phormidium uncinatum — То же, что и выше, но без фазового контраста. 200Х | |||
Формидиум sp. | |||
Формидиум sp. | |||
Диаграмма фикобилисомы Каждый диск в структуре фикобилисом представляет собой гексамерный агрегат (альфа-бета)6 6 фикобилипротеина в комплексе с линкерным белком: фикоэритрин = красный, фикоцианин = синий, аллофикоцианин = голубой. Зеленые элементы — это два реакционных центра ФСII, которые связаны с каждой фикобилисомой. | |||
Planktothrix (Oscillatoria) agardhii — С видимыми псевдовакуолями. 400-кратный фазовый контраст | |||
Plankothrix agardhii — цветение, содержащее микроцистин, в пресноводном озере Шотландии | |||
Plankothrix agardhii — прибрежные отложения цветка, содержащего микроцистин, в пресноводном шотландском озере | |||
Planktothrix rubescens . | |||
Plectonema crispatum | |||
Pleurocapsa sp. ул. ПКК 7440 | |||
Псевдоанабаена африканская | |||
Rhaboderma rubrum | |||
Raphidiopsis curvata — Смешанные с некоторыми прямыми цепочками, которые либо более Raphidiopsis , либо Cylndrospermopsis raciborskii без акинет или гетероцист. 400Х | |||
Raphidiopsis curvata — То же, что и выше, но с фазовым контрастом | |||
Рафидиопсис ср. mediterranea — может быть R. brookii . 400Х | |||
Рафидиопсис ср. средиземноморье — 100X | |||
Raphidiopsis curvata — 200X | |||
Рафидиопсис ср. | |||
Ривулярия sp. из Claxton Manor в Норфолке, Великобритания | |||
Ривулярия sp. из Claxton Manor в Норфолке, Великобритания | |||
Ривулярия sp. из Claxton Manor в Норфолке, Великобритания | |||
Ривулярия сп. из Claxton Manor в Норфолке, Великобритания | |||
Ривулярия sp. из Claxton Manor в Норфолке, Великобритания | |||
Schizothrix arenaria | |||
Schizothrix calcicola | |||
шизотрикс ленормандский | |||
Schizothrix turicensis | |||
Schizothrix viguieri | |||
Сцитонема ср. | |||
Сцитонема ракообразная | |||
Сцитонема Гофманни | |||
Scytonema javanicum | |||
Сцитонема обскурум | |||
Scytonema ocellatum | |||
Scytonema schmidtii | |||
Scytonema stuposum | |||
Сцитонема вариум | |||
Сцитонема sp. | |||
Spirulina major — фазовый контраст 400X | |||
Спирулина sp. | |||
Спирулина sp. | |||
Спирулина sp. | |||
Спирулина sp. | |||
Спирулина sp. | |||
Спирулина sp. | |||
Спирулина sp. | |||
Спирулина sp. | |||
Спирулина субсальса | |||
Спирулина sp. ( Артроспира ) культура, зараженная бурыми лебедями в Саут-Бенде, Индиана. | |||
Строматолиты | |||
Строматолиты | |||
Строматолиты | |||
Строматолиты | |||
Столбчатые строматолиты и тромболиты из приливного канала на острове Ли-Стокинг, Багамы | |||
Строматолит Поперечный разрез столбчатого прокариотического строматолита, выращенного в норе Калианасса . | |||
Строматолит Микрофотография тонкого среза прокариотического строматолита, показывающая чередующиеся слои плотного микрита и грубого детрита. | |||
Строматолит Микрофотография тонкого среза эукариотического строматолита, показывающая волнистые пластинки, образованные сочетанием активности эукариотических водорослей и цианобактерий. | |||
Строматолиты, обнаруженные в среднепротерозойских образованиях сланцев Хакатаи в национальном парке Гранд-Каньон. | |||
Строматолиты, обнаруженные в среднепротерозойских образованиях сланцев Хакатаи в национальном парке Гранд-Каньон. Крышка объектива 55 мм. | |||
Строматолиты, обнаруженные в среднепротерозойских образованиях сланцев Хакатаи в национальном парке Гранд-Каньон. Крышка объектива 55 мм. | |||
Строматолиты, обнаруженные в среднепротерозойских образованиях сланцев Хакатаи в национальном парке Гранд-Каньон. | |||
Разрез строматолита | |||
Замороженные строматолиты, Шаркс Бэй, Австралия | |||
Замороженные строматолиты, Шаркс-Бей, Австралия | |||
Мат из замороженного строматолита, Шаркс-Бей, Австралия | |||
Замороженный строматолит, Шаркс Бэй, Австралия | |||
Замороженные строматолиты, Шаркс Бэй, Австралия | |||
Живые строматолиты, озеро Салда, Турция | |||
Живые строматолиты, озеро Салда, Турция | |||
Живые строматолиты, озеро Салда, Турция | |||
Живые строматолиты, озеро Салда, Турция | |||
Живые строматолиты, озеро Салда, Турция | |||
Живые строматолиты, озеро Салда, Турция | |||
Живые строматолиты, озеро Салда, Турция | |||
Живые строматолиты, озеро Салда, Турция | |||
Строматоиды, дно высохшего озера у Агейского моря | |||
Строматоиды, высохшее дно озера у Агейского моря | |||
Строматоиды, дно высохшего озера у Агейского моря | |||
Symploca muscorum | |||
Synechocystis buzasii | |||
Synechocystis sp. | |||
Тромболитовые головки сильно заросли на острове Ли-Стокинг, Багамы | |||
Тромболит современного происхождения из пролива Ли Стокинг, Багамы. Бар = 1 см. | |||
Микрофотография тромболита, тонкий срез, показывающий границу между микритовым сгустком и карманом осадка, богатым детритом | |||
Tolypothrix bouteillei | |||
Tolypothrix byssoidea | |||
Tolypothrix campylonemoides | |||
Толипотрикс искаженный | |||
Tolypothrix phyllophila | |||
Tolypothrix sp. штамм А | |||
Tolypothrix sp. |
 вероятно 400X
вероятно 400X 200Х
200Х — (Необычный и искаженный) Тот же образец, что и выше, при 100-кратном увеличении.
— (Необычный и искаженный) Тот же образец, что и выше, при 100-кратном увеличении. Обратите внимание на сказочные прорастающие акинеты.
Обратите внимание на сказочные прорастающие акинеты. 400Х
400Х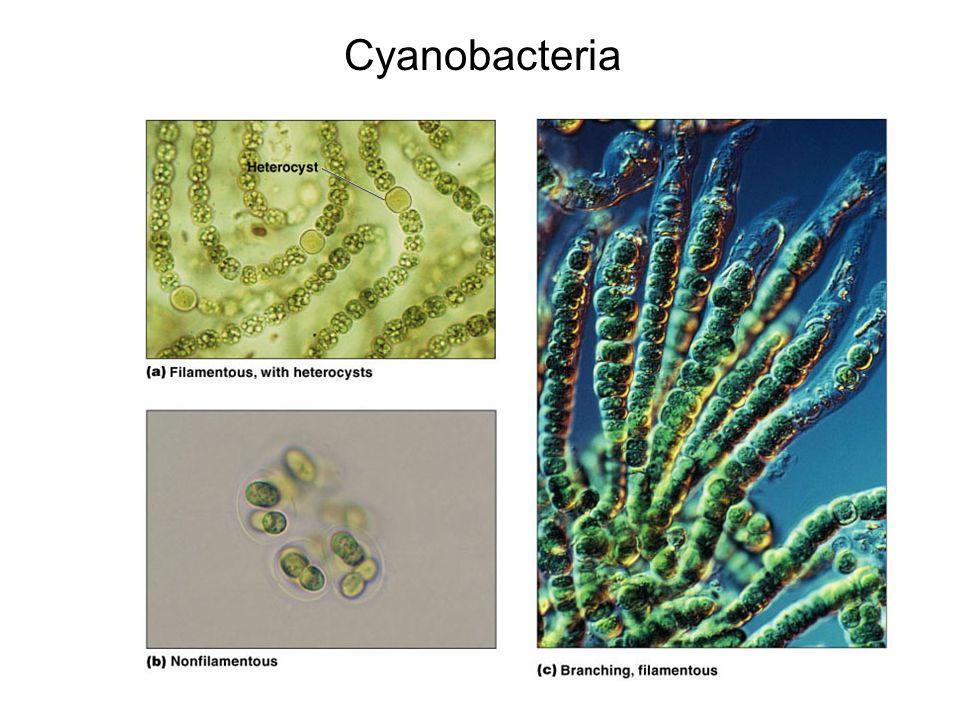 протокол.
протокол.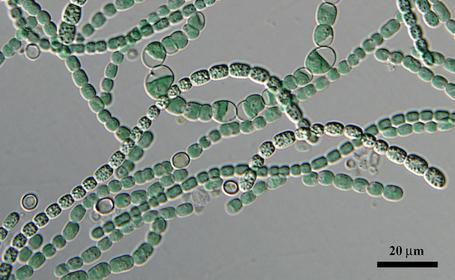 400Х
400Х

 из системы переработки целлюлозно-бумажных отходов в Бразилии
из системы переработки целлюлозно-бумажных отходов в Бразилии 400-кратный фазовый контраст
400-кратный фазовый контраст


 — (Неизвестно) Возможно биргеи . 200Х
— (Неизвестно) Возможно биргеи . 200Х 200Х
200Х 400Х
400Х 1000X
1000X — Становление сливным. 200X
— Становление сливным. 200X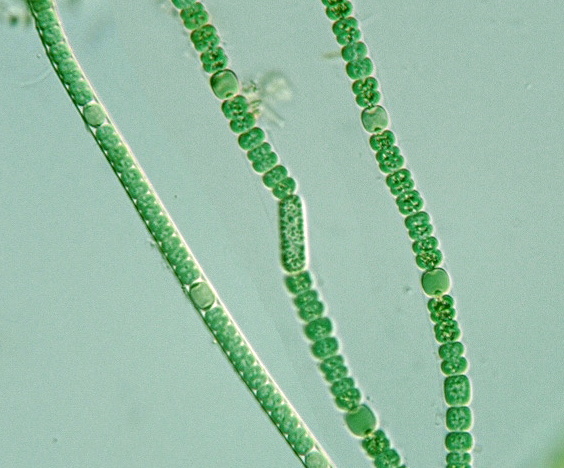
 200X. См. Mystery Bug A.
200X. См. Mystery Bug A. цветение на месте из подводного иллюминатора в Балтийском море
цветение на месте из подводного иллюминатора в Балтийском море , 130X
, 130X
 200X
200X вместе с микроколонией Synechocystis . Обратите внимание на многочисленные бактерии на заднем плане.
вместе с микроколонией Synechocystis . Обратите внимание на многочисленные бактерии на заднем плане. 200X
200X 100X.
100X. mediterranea — отчетливо видны псевдовакуоли. 400-кратный фазовый контраст
mediterranea — отчетливо видны псевдовакуоли. 400-кратный фазовый контраст Ракообразные
Ракообразные