Учебный сайт Полины Байкузиной. В форма днк
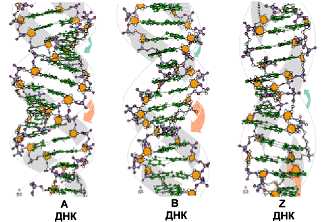
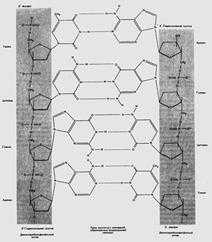
Изображения молекул ДНК форм A, B и Z
В ДНК при помощи комбинаций четырех различных мономерных молекул — нуклеотидов, соединенных в длинную цепь, записана информация о строении белков организма. Две цепи дезоксирибонуклеотидов взаимодействуют друг с другом по определенному правилу — принципу комплементарности, (аденин связывается с тимином, а гуанин с цитозином) с образованием протяженной двойной спирали. В ядре одной средней человеческой клетки в объеме 40 кубических микрометров упаковано 2 метра ДНК (1).
Строение ДНК в разных формах
Молекула ДНК может в зависимости от условий существовать в разных формах. Наиболее распространой является В-форма. В этой форме находится основная часть ДНК в клетках. При такой организации плоскости азотистых оснований практически перпендикулярны оси двойной спирали, и каждая пара повёрнута относительно предыдущих на 36o. На один виток спирали приходится примерно 10 нуклеотидных пар (9,7 и 10,6 в различных кристаллах)(2), а длина составляет 3,4 нм.
Диаметр двойной спирали В-форме (расстояние между атомами фосфора одной комплементарной пары) составляет 2 нм, причём пуриновые нуклеотиды занимают 3/5 этого расстояния, а пиримидиновые — 2/5. Глубина большой бороздки составляет 0,85 нм, а малой — 0,75 нм. При этом ширина большой бороздки достигает 1,2 нм, что приблизительно в два раза больше ширины малой бороздки (3).
Установлено, что понижение относительной влажности при добавлении неэлектролитов (например, этанола) способствует переходу ДНК из В формы в А-форму.Это сопровождается изменением С2’-эндо конформации молекул дезоксирибозы на С3’-эндо конформацию. При этом плоскости азотистых оснований отклоняются от оси спирали примерно на 13. На один виток в этой форме приходится чуть меньше 11 нуклеотидных пар (4).
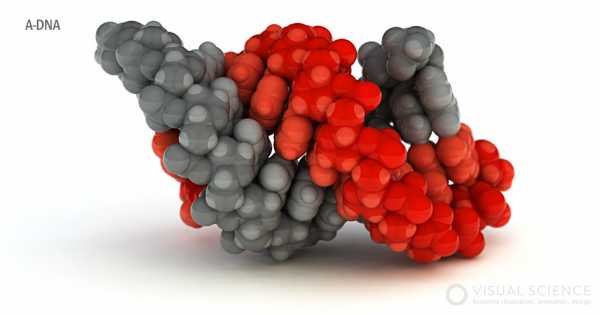
A-ДНК, вид сбоку

A-ДНК, вид с торца
Существенным отличием А-формы от В-формы является то, что в А-форме пары оснований сдвинуты к периферии спирали почти на половину её радиуса, в результате чего пространство вдоль оси оказывается пустым. Большая бороздка при этом становится глубже и уже, а малая бороздка оказывается шире и более плоской (5). ДНК может переходить в А-форму не только при пониженной относительной влажности, но и в составе гетеродуплекса с молекулами РНК, для которой А-форма оказывается более устойчивой из-за наличия дополнительной группы ОН- рибозы. Таким образом, в клетке А-форма ДНК всегда образуется во время транскрипции, обратной транскрипции и при отжиге РНК-праймеров в ходе репликации.
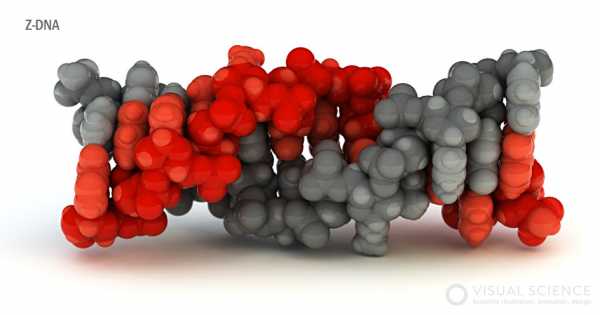
Z-ДНК, вид сбоку

Z-ДНК, вид с торца
Помимо В-формы и А-формы встречается также Z-форма (нижнее изображение) двуцепочечной ДНК. В отличие от первых двух, она представляет собой левозакрученную спираль с длиной витка 4,4 нм, на который приходится 12 нуклеотидных пар. В эту форму при понижении влажности или увеличении концентрации солей чаще всего переходят участки молекулы, богатые CpG-повторами. Наличие ДНК в Z-форме характерно для некоторых энхансеров (6, 7). Также Z-форма может образовываться за работающей РНК-полимеразой в результате отрицательной сверхспирализации молекулы.
Описаны также и другие формы организации двойной спирали ДНК — Н-форма, B’-форма, С-форма и D-форма. Однако они встречаются гораздо реже и играют менее значительную физиологическую роль.
На изображениях антипараллельные цепи окрашены серым и красным цветами. Сахарофосфатный остов имеет более темные оттенки.
Показать ссылки
visual-science.com
В-форма [ДНК] - это... Что такое В-форма [ДНК]?
ДНК C C-форма ДНК — ДНК C, C форма ДНК * ДНК C, C форма ДНК * C DNA or C helix конформация двунитчатой ДНК, при которой две нити Уотсона Крика спирали () образуют правозакрученную спиральную структуру, в которой на один виток с большой и малой бороздками приходится… … Генетика. Энциклопедический словарь
D-форма ДНК — * D форма ДНК * D form of DNA правоспиральное конформационное состояние двунитевой молекулы ДНК. Оно может проявляться только в участках ДНК, лишенных гуанина. Число пар оснований на виток в такой молекуле меньше, чем в более распространенных… … Генетика. Энциклопедический словарь
А-форма ДНК — * А форма ДНК * A form DNA см … Генетика. Энциклопедический словарь
В-форма [ДНК] — Правоспиральное конформационное состояние молекулы ДНК, существующее при высокой относительной влажности (>92 %) и в растворах с низкой ионной силой; полагают, что в живых клетках практически вся ДНК существует именно в В ф. (в 80 х гг. было… … Справочник технического переводчика
D-форма [ДНК] — D form D форма [ДНК]. Правоспиральное конформационное состояние двунитевой молекулы ДНК, может появляться только в лишенных гуанина молекулах ДНК; число пар оснований на виток меньше, чем в более распространенных формах А, В и С <A form, B… … Молекулярная биология и генетика. Толковый словарь.
Z-форма [ДНК] — Z form, zig zag DNA Z форма [ДНК]. Левоспиральное (в отличие от всех других A <A form>, B <B form>, C <C form>, D форм <D form>) конформационное состояние двухцепочечной молекулы ДНК, существующее при очень высоких… … Молекулярная биология и генетика. Толковый словарь.
С-форма [ДНК] — C form С форма [ДНК]. Правоспиральное конформационное состояние двуцепочечной молекулы ДНК, образующееся при влажности 66% и в присутствии ионов лития; число пар оснований на 1 виток 9,1/3, расстояние между парами оснований 3,32 , угол вращения… … Молекулярная биология и генетика. Толковый словарь.
А-форма [ДНК] — Правоспиральное конформационное состояние двухцепочечной молекулы ДНК, возникающее при 75 % ной влажности и в присутствии ионов калия, натрия или цезия, число пар оснований на виток 11, расстояние между парами оснований 2,56 , угол вращения между … Справочник технического переводчика
С-форма [ДНК] — Правоспиральное конформационное состояние двуцепочечной молекулы ДНК, образующееся при влажности 66 % и в присутствии ионов лития; число пар оснований на 1 виток 9,1/3, расстояние между парами оснований 3,32 , угол вращения между соседними парами … Справочник технического переводчика
Z-форма ДНК — DNR Z forma statusas T sritis chemija apibrėžtis Kairiojo sukimo dvivijė DNR spiralė, išsidėsčiusi zigzagu. atitikmenys: angl. DNA Z form rus. Z форма ДНК … Chemijos terminų aiškinamasis žodynas
dic.academic.ru
Формы ДНК, структура и синтез
Дезоксирибонуклеиновая кислота – ДНК – служит носителем наследственной информации, передаваемой живыми организмами следующим поколениям, и матрицей для строительства белков и различных регуляторных факторов, потребных организму в процессах роста и жизнедеятельности. В этой статье мы остановимся на том, какими бывают наиболее распространенные формы структуры ДНК. Также мы обратим внимание на то, как строятся эти формы и в каком виде ДНК пребывает внутри живой клетки.
Уровни организации молекулы ДНК
Различают четыре уровня, определяющих строение и морфологию этой гигантской молекулы:
- Первичный уровень, или структура – это порядок нуклеотидов в цепи.
- Вторичная структура представляет собой знаменитую «двойную спираль». Устоялось именно это словосочетание, хотя на самом деле подобная структура напоминает винт.
- Третичная структура образуется вследствие того, что между отдельными участками двухцепочечной закрученной нити ДНК возникают слабые водородные связи, придающие молекуле сложную пространственную конформацию.
- Четвертичная структура – это уже сложный комплекс ДНК с некоторыми белками и РНК. В такой конфигурации ДНК упакована в хромосомы в ядре клетки.

Первичная структура: компоненты ДНК
Блоками, из которых строится макромолекула дезоксирибонуклеиновой кислоты, являются нуклеотиды, представляющие собой соединения, в состав каждого из которых входят:
- азотистое основание – аденин, гуанин, тимин или цитозин. Аденин и гуанин относятся к группе пуриновых оснований, цитозин и тимин – пиримидиновых;
- пятиуглеродный моносахарид дезоксирибоза;
- остаток ортофосфорной кислоты.
При образовании полинуклеотидной цепочки важную роль играет порядок групп, образуемых атомами углерода в кольцевой молекуле сахара. Фосфатный остаток в нуклеотиде соединен с 5’-группой (читается «пять прайм») дезоксирибозы, то есть с пятым атомом углерода. Наращивание цепочки происходит путем присоединения к свободной 3’-группе дезоксирибозы фосфатного остатка следующего нуклеотида.
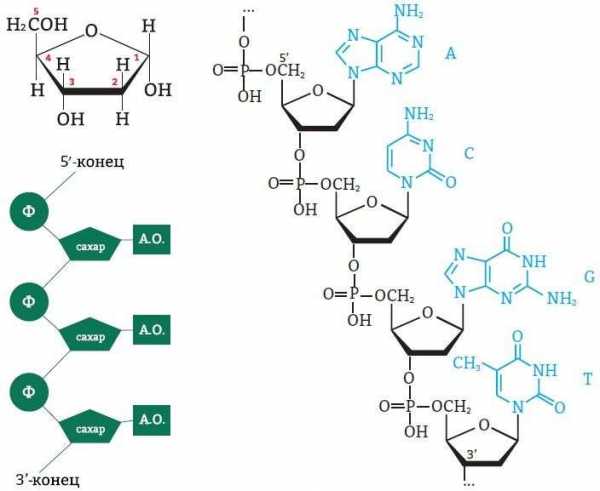
Таким образом, первичная структура ДНК в форме полинуклеотидной цепи – имеет 3’- и 5’-концы. Это свойство молекулы ДНК называется полярностью: синтез цепи может идти только в одном направлении.
Образование вторичной структуры
Следующий шаг в структурной организации ДНК базируется на принципе комплементарности азотистых оснований – их способности попарно соединяться друг с другом посредством водородных связей. Комплементарность – взаимное соответствие – возникает по той причине, что аденин и тимин образуют двойную связь, а гуанин и цитозин – тройную. Поэтому при формировании двойной цепи эти основания встают друг напротив друга, образуя соответствующие пары.
Полинуклеотидные последовательности располагаются во вторичной структуре антипараллельно. Так, если одна из цепочек имеет вид 3’ – АГГЦАТАА – 5’, то противоположная будет выглядеть следующим образом: 3’ – ТТАТГЦЦТ – 5’.
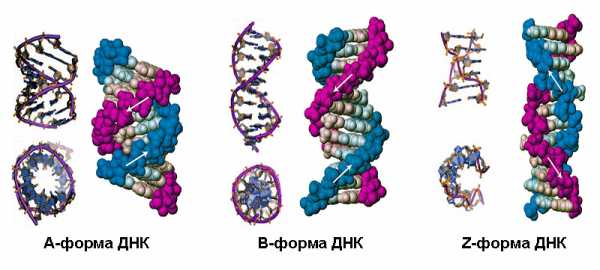
При образовании молекулы ДНК происходит закручивание сдвоенной полинуклеотидной цепи, причем от концентрации солей, от водонасыщенности, от строения самой макромолекулы зависит, какие формы может принимать ДНК на данной структурной ступени. Известно несколько таких форм, обозначаемых латинскими буквами A, B, C, D, E, Z.

Конфигурации C, D и E не встречены в живой природе и наблюдались только в лабораторных условиях. Мы рассмотрим основные формы ДНК: так называемые канонические A и B, а также конфигурацию Z.
А-ДНК – сухая молекула
А-форма – это правый винт с 11 комплементарными парами оснований в каждом витке. Диаметр его составляет 2,3 нм, а длина одного витка спирали – 2,5 нм. Плоскости, образуемые спаренными основаниями, имеют наклон 20° по отношению к оси молекулы. Соседние нуклеотиды расположены в цепочках компактно – между ними всего 0,23 нм.
Такая форма ДНК возникает при низкой гидратации и при повышенной ионной концентрации натрия и калия. Она характерна для процессов, в которых ДНК образует комплекс с РНК, поскольку последняя не способна принимать иные формы. Кроме того, А-форма весьма устойчива к ультрафиолетовому облучению. В этой конфигурации дезоксирибонуклеиновая кислота содержится в грибных спорах.
Влажная B-ДНК
При малом содержании солей и высокой степени гидратации, то есть в нормальных физиологических условиях, ДНК принимает свою главную форму B. Природные молекулы существуют, как правило, в В-форме. Именно она лежит в основе классической модели Уотсона-Крика и чаще всего изображается на иллюстрациях.
Данной форме (она также правозакрученная) свойственна меньшая компактность размещения нуклеотидов (0,33 нм) и большой шаг винта (3,3 нм). Один виток содержит 10,5 пары оснований, поворот каждой из них относительно предыдущей составляет около 36°. Плоскости пар почти перпендикулярны оси «двойной спирали». Диаметр такой сдвоенной цепочки меньше, чем у А-формы – он достигает только 2 нм.
Неканоническая Z-ДНК
В отличие от канонических ДНК, молекула типа Z представляет собой левозакрученный винт. Она самая тонкая из всех, имеет диаметр всего 1,8 нм. Витки ее длиной 4,5 нм как бы вытянуты; эта форма ДНК содержит 12 спаренных оснований на каждый виток. Расстояние между соседними нуклеотидами также достаточно велико – 0,38 нм. Так что Z-форма характеризуется наименьшей степенью скрученности.
Образуется она из конфигурации B-типа на тех участках, где в составе нуклеотидной последовательности чередуются пуриновые и пиримидиновые основания, при изменении содержания ионов в растворе. Формирование Z-ДНК связано с биологической активностью и является очень кратковременным процессом. Подобная форма нестабильна, что создает трудности при исследовании ее функций. Пока что они в точности не ясны.
Репликация ДНК и ее строение
И первичная, и вторичная структуры ДНК возникают в ходе явления, называемого репликацией – образования из материнской макромолекулы двух идентичных ей «двойных спиралей». При репликации исходная молекула расплетается, и на освободившихся одиночных цепочках происходит наращивание комплементарных оснований. Поскольку половинки ДНК антипараллельны, этот процесс протекает на них в разных направлениях: по отношению к материнским цепочкам от 3’-конца к 5’-концу, то есть новые цепочки растут в направлении 5’ → 3’. Лидирующая цепь синтезируется непрерывно в сторону репликационной вилки; на отстающей цепи синтез совершается от вилки отдельными участками (фрагменты Оказаки), которые затем сшивает между собой особый фермент – ДНК-лигаза.
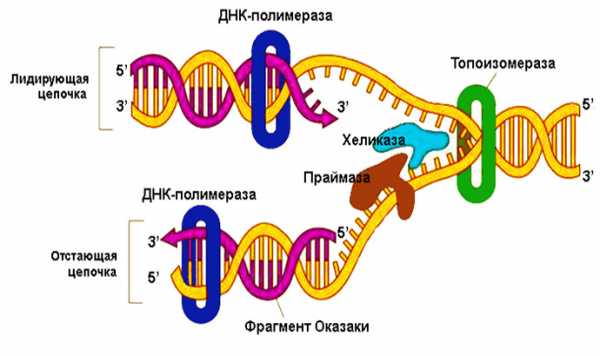
Пока продолжается синтез, уже сформированные концы дочерних молекул претерпевают винтообразное закручивание. Затем, еще до окончания репликации новорожденные молекулы начинают образовывать третичную структуру в процессе, именуемом сверхспирализацией.
Суперскрученная молекула
Сверхспирализованная форма ДНК возникает, когда двухцепочечная молекула совершает дополнительное закручивание. Оно может быть направлено по часовой стрелке (положительно) либо против (в этом случае говорят об отрицательной сверхспирализации). ДНК большинства организмов суперскручена отрицательно, то есть против основных витков «двойной спирали».
В результате образования дополнительных петель – супервитков – ДНК приобретает сложную пространственную конфигурацию. В клетках эукариот этот процесс происходит с формированием комплексов, в которых ДНК отрицательно навивается на гистоновые белковые комплексы и принимает вид нити с бусинами-нуклеосомами. Свободные участки нити называются линкерами. В поддержании суперскрученной формы молекулы ДНК принимают участие и негистоновые белки, а также неорганические соединения. Так образуется хроматин – вещество хромосом.
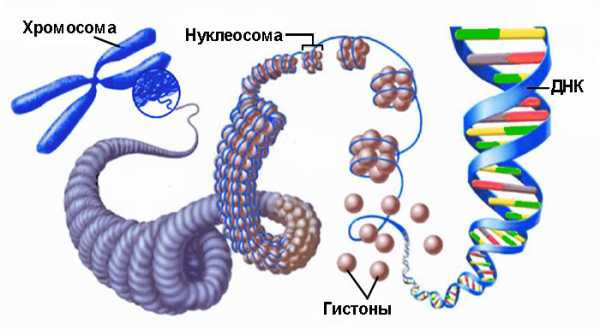
Хроматиновые нити с нуклеосомными бусинами способны к дальнейшему усложнению морфологии в процессе, называемом конденсацией хроматина.
Окончательная компактизация ДНК
В ядре форма макромолекулы дезоксирибонуклеиновой кислоты становится чрезвычайно сложной, компактизируясь в несколько этапов.
- Во-первых, нить сворачивается в особую структуру типа соленоида – хроматиновую фибриллу толщиной в 30 нм. На этом уровне ДНК, сворачиваясь, сокращает свою длину в 6-10 раз.
- Далее фибрилла при помощи специфических скэффолд-белков образует зигзагообразные петли, что уменьшает линейный размер ДНК уже в 20-30 раз.
- На следующем уровне формируются плотно упакованные петельные домены, чаще всего имеющие форму, условно названную «ламповая щетка». Они прикрепляются к внутриядерному белковому матриксу. Толщина таких структур составляет уже 700 нм, ДНК при этом укорачивается приблизительно в 200 раз.
- Последний уровень морфологической организации – хромосомный. Петельные домены уплотняются настолько, что достигается общее укорочение в 10 000 раз. Если длина растянутой молекулы – около 5 см, то после упаковки в хромосомы она уменьшается до 5 мкм.
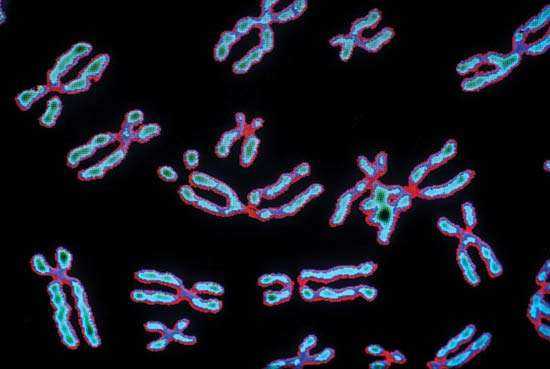
Высшего уровня усложнения формы ДНК достигает в состоянии метафазы митоза. Именно тогда она приобретает характерный облик – две хроматиды, соединенные перетяжкой-центромерой, которая обеспечивает расхождение хроматид в процессе деления. Интерфазная ДНК организована до доменного уровня и распределяется в ядре клетки без особого порядка. Таким образом, мы видим, что морфология ДНК тесно связана с различными фазами ее существования и отражает особенности функционирования этой важнейшей для жизни молекулы.
fb.ru
8. Полиморфизм вторичной структуры днк
В настоящее время известно, что пространственная структура ДНК обладает полиморфизмом, то есть она способна принимать различные конформации. Рентгеноструктурные исследования кристаллов олигонуклеотидов выявили три основных типа структур - А-, В- и Z- формы. В-ДНК - это стандартная структура, описанная Уотсоном и Криком, в которой плоскости пар оснований перпендикулярны оси двойной спирали. Геометрия её такова, что соседние пары оснований находятся друг от друга на расстоянии 0,34 нм и повернуты на 360 вокруг оси спирали. На один виток спирали приходится, следовательно, 10 пар оснований (3600/360 = 10), и шаг спирали равен 3,4 нм (10 . 0,34 нм). Диаметр двойной спирали равен примерно 20 нм. В А-ДНК плоскости пар оснований повернуты примерно на 200С от нормали к оси правой двойной спирали. На виток спирали здесь приходится 11 пар оснований. А-ДНК образуется при высушивании волокон В-ДНК. В Z-ДНК буква Z указывает на зигзагообразную форму сахарофосфатного остова ДНК в этой форме. Плоскости оснований примерно перпендикулярны оси спирали. В клетке ДНК обычно находится в В-форме, но отдельные её участки вследствие сверхспирализации могут быть в А-, Z- или даже в иной конформации.
| | Отличительные особенности форм ДНК: А ДНК - правозакрученная спираль, короткая и широкая; шаг - 2,3 А; 1 виток - 11 пар оснований. В ДНК - правозакрученная спираль, длиннее и тоньше, шаг - 3,32 А; 1 виток - 10 пар оснований. Z ДНК - левозакрученная спираль, самая длинная и самая тонкая; шаг - 3,8 А; 1 виток - 12 пар оснований |
Ни А-, ни Z- формы не могут существовать в водном растворе без дополнительных воздействий (белки или суперспирализация)
9. Механизмы хранения и передачи наследственной информации - репарация, репликация (строение репликативной вилки), транскрипция, трансляция, характеристика основных ферментов и кофакторов.
Репликация («самоудвоение») - перенос генетической информации в пределах одного класса нуклеиновых кислот, т.е. от ДНК к ДНК (происходит полное копирование информации).
Транскрипция («переписывание») – перенос информации между разными классами нуклеиновых кислот: от ДНК к РНК (от РНК к ДНК – у вирусов), т.е. происходит копирование отдельных участков. В ходе транскрипции образуются все виды РНК (м-РНК, т-РНК, р-РНК)
Трансляция – перенос генетической информации от м-РНК к белку, т.е. в пределах разных классов молекул: перевод информации с «языка» нуклеиновых кислот на «язык» полипептидной цепи.
Репарация ДНК – ограниченная репликация, исправление поврежденных участков ДНК ДНКазами, затем ДНК-полимеразы заполняют пробел и концы «сшиваются» ДНК лигазами (см. рисунок).
Репликация ДНК (воспроизведение генотипа) происходит по полуконсервативному механизму, то есть дочерняя цепь ДНК синтезируется на материнской цепи, называемой обычно матрицей. Следовательно, вновь образованные двухспиральные молекулы состоят из одной «новой» и одной «старой» цепи.
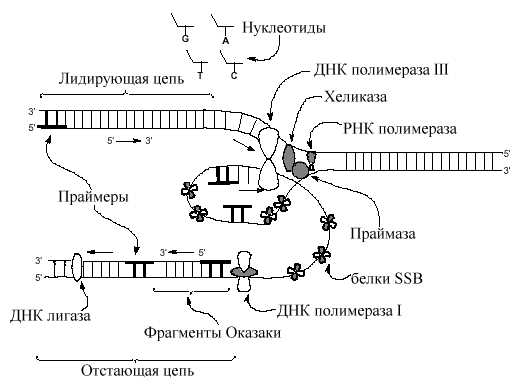
1. Все ДНК-полимеразы нуждаются для начала своей работы в предварительно синтезированных олигонуклеотидах – затравках, 3’-ОН группа которых используется для удлинения цепи. (Эти короткие отрезки, обычно, рибонуклеотидов синтезируются специальными РНК-полимеразами. По названию затравки – праймер эти полимеразы получили название праймаз). Длина праймеров составляет 10-12 нуклеотидов. Праймеры синтезируются на обеих цепях.
2. Далее праймер достраивается ДНК-полимеразой – III. Т.к. все ДНК-полимеразы формируют дочерние цепи в одном направлении 5’ 3’, то одна цепь синтезируется непрерывно (лидирующая, ведущая цепь), а другая - прерывисто (отстающая). ДНК-полимераза формирует димер, связанный с другими необходимыми в репликативной вилке белками (реплисома). Одна из цепей матричной ДНК (отстающая), временно образует петли вокруг реплисомы так, что димер ДНК-полимеразы получает возможность перемещаться по обеим цепям в одном 3' 5 ' направлении одновременно. Короткие (до 1000 пар нуклеотидов) отрезки ДНК, образующиеся на отстающей цепи, получили название фрагментов Оказаки по имени Ф. Оказаки, впервые (1968) указавшего на прерывистый характер синтеза ДНК. 3.Так как репликативная вилка довольно быстро (1000 нулеотидов в сек) продвигается по матрице, вновь синтезируемые дочерние цепи и родительские цепи матрицы сразу формируют двойные спирали ДНК. Предполагается, что только маленький отрезок матричной двойной спирали находится в одноцепочечном состоянии в данный отрезок времени.
3. Праймеры ведущей и отстающей цепей (10-12 пар нуклеотидов) удаляются ДНК полимеразой I, обладающей репарирующей функцией с одновременной заменой рибонуклеотидов дезоксирибонуклеотидами.
4. Промежутки, которые возникают между 3 '-ОН и 5 '- фосфатом, «сшиваются» лигазами ДНК, завершая тем самым процесс репликации.
| Основные ферменты и белки, участвующие в процессе репликации | |
| Белок | Функция |
| Топоизомераза I | Ослабляет суперспирализацию ДНК |
| Белок Rep Хеликаза | раскручивает ДНК в репликационной вилке |
| SSB | Связывается с одноцепочечной ДНК в репликативной вилке для предотвращения образования двойной спирали |
| Праймаза (РНК-полимераза) | Синтезирует РНК-затравку |
| ДНК-полимераза III | Полимераза, катализирующая репликацию |
| ДНК-полимеразаI | Удаляет РНК-затравку, заполняя пробел |
| ДНК лигаза | Катализирует образование 3 - 5 -фосфодиэфирной связи, соединяя фрагменты Оказаки |
| Топоизомераза II | Разделяет ДНК после репликации |
studfiles.net
Альтернативные структуры ДНК
Рассмотренную структуру ДНК обычно считают одним из несомненных фактов молекулярной биологии. Но недавно стало очевидным, что некоторые параметры классической В-формы нужно пересмотреть и даже что ДНК может образовывать другие типы двуспиральных структур.
Проблема определения точных значений параметров двойной спирали связана с тем, что все первоначальные значения были получены при изучении дифракции рентгеновских лучей на волокнах ДНК. По этим данным установлены основные характеристики, такие, как число пар оснований на виток, расстояние между соседними парами вдоль оси спирали. Однако этим методом нельзя определить положения отдельных атомов, как это делают, используя рентгеноструктурный анализ. Поэтому модель уточняют, согласуя расчетную картину дифракции рентгеновских лучей с экспериментальными данными. Теоретически на основе одних и тех же данных всегда можно построить разные модели. Дело в том, что модель удовлетворяет усредненным данным и, следовательно, в определенных областях структуры возможны вариации. Например, до сих пор мы говорили о ДНК как о длинной, жесткой двойной спирали, но мы знаем, что в действительности она должна быть свернута и плотно уложена, для того чтобы уместиться в клетке. При этом детали ее структуры могут изменяться.
Сейчас считается, что двухцепочечная ДНК-всегда двойная спираль. Однако была предложена и другая модель, хорошо согласующаяся с данными дифракции рентгеновских лучей. Согласно этой модели, две антипараллельные цепи ДНК также соединяются путем комплементарного спаривания оснований, но лежат «бок о бок», вместо того, чтобы закручиваться в непрерывную двойную спираль.
Существование ДНК в виде двойной спирали было подтверждено экспериментами, в которых прямо измеряли число пар оснований на виток. Оказалось, что их 10,4 вместо 10,0, предсказанных классической В-моделью. Это различие вызвало необходимость несколько изменить угол вращения между соседними парами оснований вдоль спирали до 34,6°, так что отрезок спирали, в пределах которого совершается полный виток на 360°, стал несколько длиннее.
Особенно важно, что значение 10,4 является средним для ДНК как целой молекулы при определенных условиях. Изменение условий или даже последовательности отдельных оснований может привести к большему или меньшему закручиванию спиральной структуры в соответствующих участках.
Действительно, методом рентгеноструктурного анализа было показано, что молекула, состоящая из 12 пар оснований, содержит 10,1 пары оснований на виток, что обеспечивается слабым сдвигом каждой пары оснований, при котором улучшаются межплоскостные (стэкинг) взаимодействия между основаниями, по сравнению с первоначальной моделью.
Ввиду таких вариаций идея о существовании единственной структуры двуспиральной ДНК сменилась представлением, допускающим наличие семейства структур, каждая из которых имеет характерный тип, но проявляет различия по главным параметрам - n (число нуклеотидов на виток) и h (расстояние между соседними повторяющимися элементами). Вариации обусловлены изменением вращения отдельных групп вокруг связей, обладающих свободой вращения. В пределах каждого семейства структур параметры могут слабо варьировать. Например, для В-ДНК значение n может составлять от 10,0 до 10,6.
В течение долгого времени были известны три структурные формы ДНК, способные к взаимопревращениям при изменении соответствующих условий. В-форма спирали, для которой Уотсон и Крик построили свою модель, характерна для волокон ДНК при очень высокой относительной влажности (92%) и в растворах низкой ионной силы. Считают, что именно в такой форме ДНК обычно находится в живой клетке.
А-форма обнаружена в волокнах ДНК при 75% влажности и нуждается в присутствии ионов натрия, калия или цезия, несущих противоположный заряд. Основания, располагавшиеся строго перпендикулярно оси спирали в В-форме, в А-форме наклонены по отношению к оси спирали, и их число на виток больше. А-форма интересна с биологической точки зрения, так как ее конформация очевидно, близка к структуре гибридов ДНК-РНК и двуспиральных участков РНК (обсуждается далее). Причина этого заключается в том, что 2'-гидроксильная группа мешает ДНК принять В-форму.
С-форма образуется, когда ДНК находится при 66% влажности в присутствии ионов лития. У нее меньше пар оснований на виток, чем у В-ДНК.
В этих трех формах могут находиться все ДНК независимо от их нуклеотидной последовательности. Следующие формы характерны только для молекул ДНК с определенными особенностями в составе пар оснований.
D-форма и Е-форма (возможно, крайние варианты одной и той же формы) имеют наименьшее число пар на виток (8 и 7,5) и обнаружены только в определенных молекулах ДНК, не содержащих гуанина.
Z-форма представляет собой наиболее резкий контраст с классическим структурным семейством. Эта форма левоспиральная, тогда как все остальные - правоспиральные. Она имеет наибольшее число пар на виток, т.е. менее скрученная и более тонкая. Свое название форма получила из-за зигзагообразной (zigzag) линии, которую образует сахарофосфатный остов вдоль спирали. В отличие от этого остов В-формы ДНК образует плавно изгибающуюся линию, что хорошо видно на рис., где сравниваются В- и Z-формы ДНК.
Z-форма двойной спирали найдена в полимерах, образованных чередующимися пурин-пиримидиновыми последовательностями. Два исследованных полимера состояли из многократно повторенных динуклеотидных пар: poly-d (GC/CG) и poly-d (AC/TG). Буква «d» указывает, что это дезоксиформы, т.е. что это ДНК, а не РНК. Верхний и нижний ряды букв соответствуют двум цепям ДНК.
Z-форма существует только при очень высоких концентрациях соли (что объясняется необходимостью противостоять повышенному электростатическому отталкиванию между нуклеотидами, сжатыми из-за уменьшения диаметра двойной спирали Z-ДНК).
Вполне возможно, что Z-форма ДНК имеет определенное биологическое значение. Об этом свидетельствуют следующие данные: замена остатка С в полимере poly-d(GC/CG) и poly-d на 5-метилцитозин значительно повышает стабильность Z-ДНК при низкой концентрации соли. Единственное различие между 5-Ме-С и С заключается в наличии метильной группы при С-5. Эта модификация цитозина происходит in vivo в результате метилирования динуклеотидной последовательности GC/CG в некоторых участках ДНК. Отсюда вытекает, что Z-ДНК может существовать in vivo при подходящих условиях, т.е. когда последовательность соответствующего состава окажется в условиях, способствующих образованию Z-формы.
До сих пор мы рассматривали ДНК как структуру совершенно изолированную. В действительности она связана с белками, которые могут оказывать значительное влияние на возможность перехода из В-формы в Z-форму. Например, ДНК, связанная с гистонами (основные хромосомные белки эукариотического ядра), не переходит из одной формы в другую в тех условиях, когда это наблюдают у свободной ДНК. Таким образом, одним из условий, необходимых для образования Z-ДНК in vivo, по-видимому, является присутствие особых белков, стабилизирующих ее структуру.
Является ли Z-ДНК единственной левосторонней формой двойной спирали? Модели для левосторонних вариантов традиционных А-, В- и С-форм построены так, что, возможно, они и существуют. Следует помнить, что все эти формы сохраняют самую существенную в биологическом смысле особенность двойной спирали: комплементарность спаривания оснований, определяющую специфичность соединения цепей.
При построении моделей каждую форму ДНК рассматривают независимо как конструкцию, в которой находится вся молекула в определенных условиях. Но вряд ли это действительно имеет место in vivo. Большинство клеточной ДНК находится, по-видимому, в В-форме с определенными модификациями параметров спирали, варьирующими локально. И только отдельные короткие участки спирали переходят в другие формы.
А-ДНК В -ДНК Z-ДНК
Рис. Нуклеиновые кислоты могут образовывать несколько типов двойной спирали.
А-ДНК В-ДНК Z-ДНК
Таким образом, ДНК может существовать в виде структурных семейств нескольких типов, в зависимости от концентрации ионов и нуклеотидного состава молекулы. Общие свойства этих форм суммированы в табл. 1.
| Тип спирали | Число пар оснований на виток | Угол вращения1 одной пары, градусы | Расстояние между парами оснований, А | Диаметр спирали, А |
| А | + 32,7 | 2,56 | ||
| В | + 36,0 | 3,38 | ||
| С | 9, 3 | + 38,6 | 3,32 | |
| Z | -30,0 | 3,71 |
1 Угол вращения показан знаком ( + ) в случае правосторонней спирали и знаком (—) в случае левосторонней спирали.
Репликация ДНК
Способность к репликации (удвоению) ДНК определяет ее уникальные свойства по хранению и передаче наследственной информации.
Принцип репликации ДНК состоит в том, что каждая из двух полинуклеотидных нитей молекулы ДНК служит в качестве программы (матрицы) для синтеза новой (комплементарной) нити. В результате на основе одной двухцепочечной молекулы образуются две одинаковые двухцепочечные молекулы, в каждой из которых одна цепочка является старой, а другая - новой (вновь синтезированной). Такой принцип репликации ДНК был назван полуконсервативным (рис.).
Рис. Принцип полуконсервативной репликации ДНК
В соответствии с этим принципом нуклеотидная последовательность матричной (родительской) нити считывается в направлении 3'→ 5', тогда как синтез новой (дочерней) нити идет в направлении 5' → 3'. Поскольку две комплементарные цепочки родительской молекулы ДНК являются антипараллельными, то синтез новой полинуклеотидной цепочки на каждой из них идет в противоположном направлении.
Механизм репликации ДНК является достаточно сложным и, различается в случае организмов, содержащих относительно небольшие по размерам молекулы ДНК в замкнутой (кольцевой) форме (многие вирусы и бактерии), и эукариот, клетки которых имеют молекулы огромных размеров, находящиеся в линейной (незамкнутой) форме.
У прокариотов кольцевая молекула ДНК представляет собой одну структурную единицу репликации (репликон), имеющую единственную точку начала (инициации) репликации (О-пункт, coстоящий примерно из 300 нуклеотидов), в которой начинается процесс расхождения (расплетания) двух нитей родительской молекулы и матричного синтеза комплементарных копий (реплик) дочерней ДНК.
Похожие статьи:
poznayka.org
Строение, формы и свойства ДНК
Развитие генетики на современном этапе связано с расшифровкой молекулярных основ наследственности. Все основные работы в этом направлении выполнены примерно за 20 лет. Расшифровка генетического кода позволила понять структуру и функции гена, а также изучить многие вопросы влияния гена на процессы онтогенеза. Вопросам реализации генетической информации у высших организмов при их развитии в онтогенезе уделяется особо большое внимание в современной молекулярной генетике. Все дело в том, что общие основы молекулярной генетики были заложены при изучении вирусов и бактерий, у которых генетическая структура, в сравнении с высшими организмами, устроена более просто.
ДНК была открыта Иоганном Фридрихом Мишером в 1869 году. Из остатков клеток, содержащихся в гное, он выделил вещество, в состав которого входят азот и фосфор. Вначале новое вещество получило название нуклеин, а позже, когда Мишер определил, что это вещество обладает кислотными свойствами, вещество получило название нуклеиновая кислота. Биологическая функция новооткрытого вещества была неясна, и долгое время ДНК считалась запасником фосфора в организме. Более того, даже в начале XX века многие биологи считали, что ДНК не имеет никакого отношения к передаче информации, поскольку строение молекулы, по их мнению, было слишком однообразным и не могло содержать закодированную информацию.
Первым, кто предположил, что генетическая информация заключена в хромосомах, был русский ученый Н. К. Кольцов (1927 г.). Однако он считал, что функцию хранителей наследственной информации выполняют специальные белковые молекулы, способные к автосинтезу. Кольцов был близок к решению проблемы генетического кодирования, но тщательные исследования показали, что информация заключена не на белковой молекуле, а на дезоксирибонуклеиновой кислоте (ДНК), которая представляет собой высокополимерное соединение, способное к автосинтезу.
Экспериментальные доказательства роли ДНК в передаче наследственной информации получили Освальд Эвери, Колин Мак-Леон и Маклин Мак-Карти (1944 г.). Они провели трансформацию, т.е. передачу наследственных признаков от одной группы бактерий к другой. В ходе эксперимента безвредная группа бактерий приобрела патогенные свойства, в результате добавления в неё мёртвых болезнетворных бактерий. Было показано, что за вновь приобретенные признаки отвечают выделенные из пневмококков ДНК.
Экспериментальные доказательства роли нуклеиновых кислот в передаче признаков позже получили американские учёные Алфред Херши и Марта Чейз на бактериофагах (эксперименты Херши-Чейз). Они вводили радиоактивные изотопы в белки и ДНК бактериофагов, и показали, что в заражённую клетку передаётся только нуклеиновая кислота фага, а новое поколение фага содержит такие же белки и нуклеиновую кислоту, как исходный фаг.
Но точное строение ДНК, как и способ передачи наследственной информации, вплоть до 50-х годов XX века оставалось неизвестным. Хотя и было доподлинно известно, что ДНК состоит из нескольких цепочек, состоящих из нуклеотидов, никто не знал точно, сколько этих цепочек и как они соединены.
Структура двойной спирали ДНК была предложена Френсисом Криком и Джеймсом Уотсоном в 1953 году на основании рентгеноструктурных данных, полученных Морисом Уилкинсом и Розалинд Франклин. Позже предложенная Уотсоном и Криком модель строения ДНК была доказана, а их работа отмечена Нобелевской премии по медицине и физиологии в 1962 г. Среди лауреатов не было скончавшейся к тому времени Розалинды Франклин, так как премия не присуждается посмертно.
Дальнейшие работы были связаны с изучением принципа записи информации о строении белков на нуклеиновой кислоте, а также механизмов передачи этой информации к месту синтеза белка и следующему поколению при делении клетки. В 1961 году М. В. Ниренберг, Дж. Маттей , С. Очоа расшифровали и экспериментально подтвердили структуру генетического кода, закончив его исследование к 1964 году.
По современным представлениям, дезоксирибонуклеиновая кислота (ДНК) - макромолекула, обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. ДНК является своеобразной матрицей, на которой синтезируется информационная или матричная рибонуклеиновая кислота (м-РНК).
Локализация ДНК у организмов достаточно многообразна. У эукариотов ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах).
В клетках прокариотических организмов кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. И у прокариот и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами. Кроме того, одно- или двухцепочечные молекулы ДНК могут образовывать геном ДНК-содержащих вирусов.
При характеристике величины ДНК организмов используются параметры генома. Геном – это общее содержание ДНК в гаплоидном ядре. Измерения размера генома приводятся в дальтонах, парах нуклеотидов (п. н.) или пикограммах (пг). Соотношение между этими единицами измерения следующие:
1 пг = 10–9 мг = 0,6х1012 дальтон = 0,9х109 п. н.
В гаплоидном геноме человека содержится около 3,2 млрд. п. н., что равно 3,5 пг ДНК. Таким образом, в ядре одной клетки человека содержится около 7 пг ДНК. Если учесть, что средний вес клетки человека равен примерно 1000 пг, то легко рассчитать, что ДНК составляет менее 1% от веса клетки. И, тем не менее, чтобы воспроизвести самым мелким шрифтом ту огромную информацию, которая содержится в молекулах ДНК одной нашей клетки, понадобилось бы тысяча книг по 1000 страниц в каждой. Размеры генома не говорят об уровне организации его обладателя. Например, у саламандры и лилии длина молекул ДНК, содержащихся в одной клетке, в тридцать раз больше, чем у человека.
Число отдельных молекул ДНК в клетке равно числу хромосом. Длина такой молекулы в наибольшей по размеру хромосоме 1 человека составляет около 8 см. Подобных гигантских полимеров пока не выявлено ни в природе, ни среди искусственно синтезированных химических соединений. У человека длина всех молекул ДНК, содержащихся во всех хромосомах одной клетке, составляет примерно 2 метра. Следовательно, длина молекул ДНК в миллиард раз больше их толщины. Так как организм взрослого человека состоит примерно из 5х1013 – 1014 клеток, то общая длина всех молекул ДНК в организме равна 1010 км (это почти в тысячу раз больше расстояния от Земли до Солнца).
С химической точки зрения ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Другая известная нуклеиновая кислота – РНК, устроена аналогичным образом. Отличие состоит в том, что нуклеотиды РНК включают сахар рибозу.
В ДНК встречается четыре вида азотистых оснований (аденин, гуанин, тимин и цитозин). В РНК также встречается четыре вида азотистых оснований. Но вместо тимина присутствует урацил.
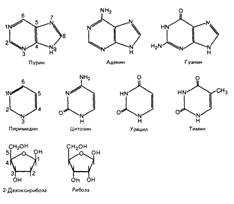
Азотистые основания представлены двумя группами: пиримидиновыми и пуриновыми. Основой пиримидинов является шестичленное пиримидиновое кольцо. Пурины имеют в своей структуре два слитых кольца- шестичленное и пятичленное.
Образование химических связей в нуклеотиде между его составляющими и отдельными нуклеотидами в полинуклеотидной цепи осуществляется через молекулу сахара. Поскольку в составе молекулы пентозы имеется 5 атомов углерода, то каждый из них можно пронумеровать индексом от 1' до 5'. В каждом нуклеотиде присоединение азотистого основания происходит к первому углеродному атому (1') пентозы с помощью β-гликозидной связи. Соединение, состоящее из углевода (пентозы) и азотистого основания, называется нуклеозидом (рис. ).
После присоединения к пятому углеродному атому остатка фосфорной кислоты образуется нуклеотид.
Название нуклеотида является производным от названия соответствующего основания. Нуклеотиды и азотистые основания обозначаются заглавными буквами.
Основание Нуклеозид Нуклеотиды (и принятые сокращения)
Аденин Аденозин Адениловая кислота = AMP (или dAMP)Гуанин Гуанозин Гуаниловая кислота = GMP (или dGMP)Цитозин Цитидин Цитидиловая кислота = СМР (или dCMP)Тимин Тимидин Тимидиловая кислота = ТМР (или dTMР)Урацил Уридин Уридиловая кислота = UMP (или dUMP)
ДНК имеет три уровня организации.
Первичная структура представлена полинуклеотидной цепью, скелетную основу которой составляют чередующиеся сахарные и фосфатные группы, связанные 3' - 5' – фосфодиэфирными ковалентными связями. Фосфоэфирные связи образуются между третьим и пятым атомами углерода соседних молекул дезоксирибозы в результате взаимодействия между 3'-гидроксильной (3'—ОН) группой одной молекулы дезоксирибозы и 5'-фосфатной группой (5'—РО3) другой. Этот остов имеет боковые группы, представленные азотистыми основаниями. Асимметричные концы цепи ДНК называются 3' (три прим) и 5' (пять прим). Полярность цепи играет важную роль при синтезе ДНК (удлинение цепи возможно только путём присоединения новых нуклеотидов к свободному 3'-концу).
У большинства организмов (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК образует вторичную структуру, представленную двумя полинуклеотидными цепями, в которых азотистые основания ориентированы друг к другу. Подобно тому, как в винтовой лестнице сбоку можно увидеть ступеньки, на двойной спирали ДНК в промежутках между фосфатным остовом молекулы можно видеть рёбра оснований, кольца которых расположены в плоскости, перпендикулярной по отношению к продольной оси макромолекулы. Эта двухцепочечная молекула спирализована. Именно, благодаря такому строению, молекула ДНК получила название «двойной спирали».
Связывание нуклеотидов двух нитей ДНК осуществляется за счет водородных связей. Азотистые основания одной из цепей соединены с азотистыми основаниями другой в строго определенном порядке согласно принципу комплементарности: аденин соединяется только с тимином, гуанин — только с цитозином. Разные пары оснований образуют разное количество водородных связей. АТ связаны двумя, ГЦ — тремя водородными связями.
Последовательность пар нуклеотидов в ДНК нерегулярна, но сами пары уложены в молекуле как в кристалле. Это дало основание характеризовать молекулу ДНК как линейный апериодический кристалл.
При определении строения ДНК важную роль сыграло наблюдение, которое известно, как «правило Чаргаффа», согласно которым в каждой молекуле ДНК соблюдаются строгие соотношения, связывающие между собой количество азотистых оснований разных типов. Согласно правилам:
1.Сумма нуклеотидов, содержащих пуриновые основания, равна сумме нуклеотидов, содержащих пиримидиновые основания (А + Г = Т + Ц).
2.Независимо от действительного количества каждого основания относительное содержание Г всегда было равно относительному содержанию Ц, а содержание А - содержанию Т. Таким образом, любая ДНК может быть охарактеризована по ее составу как отношение (Г + Ц)/(А + Т), которое, варьируя от 26 до 74%, остается характерным для каждого вида.
Основные характеристики вторичной структуры ДНК:
1. Молекула ДНК образована двумя правозакрученными полинуклеотидными цепями, т.е. если смотреть вдоль оси спирали, повороты следуют по часовой стрелке. Ширина двойной спирали составляет от 22 до 24 Å, или 2,2 - 2,4 нм, длина каждого нуклеотида 3,3 Å (0,33 нм), один поворот спирали состоит из 10 пар оснований. Каждая пара оснований повернута на 36º вокруг оси спирали относительно следующей пары оснований. Таким образом, 10 пар оснований составляют полный оборот в 360°. Две цепи, закручиваясь друг относительно друга, образуют двойную спираль, в которой имеется две бороздки - малая бороздка (около 12 Å шириной) и большая бороздка (около 22 Å шириной). Белки, например, факторы транскрипции, которые присоединяются к определённым последовательностям в двухцепочечной ДНК, обычно взаимодействуют с краями оснований в большой бороздке, где те более доступны. Такая форма ДНК встречается в природе наиболее часто и она получила название В-форма.
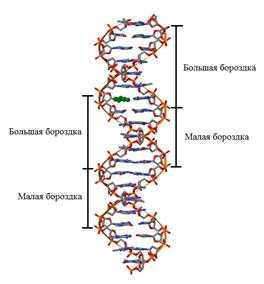
Рис. Модель вторичной структуры ДНК Уотсона Крика. Две цепи ДНК образуют двойную спираль.
Цепи объединены в результате закручивания одной цепи вокруг другой по общей оси. Из-за противоположной последовательности атомов в цепях обе цепи инвертированы относительно друг друга, т.е. направления от 3'-конца к 5'-концу в двух цепях, из которых состоит молекула ДНК, противоположны (цепи «антипараллельны» друг другу).
5΄ фосфат 3΄ гидроксильная
группа
3΄ гидроксильная 5΄ фосфат
группа
Водородные связи между основаниями двух нитей могут разрываться. Разрыв водородных связей может происходить под воздействием различных факторов, что приводит к денатурации молекулы. Например, нагревание ДНК до 950 С вызывает разрушение водородных связей. Это явление называется плавлением ДНК.
Цепочки двойной спирали могут расходиться как замок-молния и под действием ферментов (хеликазы). Причем, на разрыв ГЦ требуется больше энергии, чем на разрыв пары АТ. Процент ГЦ пар и длина молекулы ДНК определяют количество энергии, необходимой для диссоциации цепей: длинные молекулы ДНК с большим содержанием ГЦ более тугоплавки. Поэтому, части молекул ДНК, которые из-за их функций должны быть легко разделяемы, например, последовательности в бактериальных промоторах, обычно содержат большое количество А и Т.
Третичная структура ДНК определяется трехмерной пространственной организацией молекул и зависит от различных факторов.
Похожие статьи:
poznayka.org
A- и В- формы ДНК. Структура РНК
A- и В- формы ДНК. Структура РНКЗадание 1. В этом упражнении требовалось построить модели структур A-, B- и Z-формы ДНК с помощью программы fiber пакета 3DNA. Пакет 3DNA один из популярных пакетов программ для анализа и простейшего моделирования структур нуклеиновых кислот. Были построены два дуплекса ДНК, последовательность одной из нитей которых представляет собой 5 раз повторенную последовательность "gatc". Z-форма представлена 5 раз повторенной парой G-C (программа fiber позволяет строить только такую Z-форму ДНК). Структуры дуплексов сохранены в файлах A-форма, B-форма, Z-форма. Задание 2. Упражнение 1. Упражнение 2. В данном упражнении требовалось получить структуры тРНК и ДНК-белкового комплекса в формате PDB с сайта PDB. Были получены файлы со структурами тройного комплекса дрожжевой тирозил-трнк синтетазы и engrailed homeodomain-DNA. Упражнение 3. В данном упражнении требовалось проверить полученные структуры РНК и ДНК на наличие разрывов. На рис.1 изображена структура тРНК. В структуре есть разрывы, которые делят нуклеиновую кислоту на 6 фрагментов. Возможно, что структура на самом деле содержит несколько фрагментов. На рис.2 для более удобного изображения показана ленточная модель тРНК. Рис.1. Изображение тРНК Рис.2. Ленточная модель тРНК На рис.3 изображена структура ДНК. В молекуле разрывов не обнаружено. Рис.3. Изображение ДНК.Сохраненные координаты атомов ДНК и РНК. Задание 3. Упражнение 1. В данном упражнении необходимо было определить большую и малую бороздки ДНК. Для этого была взята структура B-формы ДНК. На рис.4 изображена молекула ДНК с отмеченными большой и малой бороздками. Рис.4. Изображение большой и малой бороздок в молекуле В-ДНК.Был выбран остаток тиминового нуклеотида под номером 11. При помощи программы Jmol нужно было определить какие атомы основания явно обращены в сторону большой бороздки, а какие в сторону малой. С помощью программы ChemSketch было получено изображение тимина (рис.5). Красным цветом отмечены атомы, обращенные в сторону большой бороздки, синим - в сторону малой бороздки. Рис.5. Изображение тимина. Красным цветом обозначены атомы, обращенные в сторону большой бороздки, синим - в сторону малой бороздки.
Упражнение 2. Сравнение основных спиральных параметров разных форм ДНК.
Как видно из таблицы, в А-форме ДНК большая бороздка по ширине меньше, чем малая, но большая бороздка глубже. Упражнение 3. Сравнение торсионных углов в структурах А- и В-форм.
Как видно из таблицы, значения углов A- и В-форм не совпадают со значениями, приведенными в презентации. Задание 4. Определение параметров структур нуклеиновых кислот с помощью программ пакета 3DNA. Т.к.пакет 3DNA работает только со старым форматом PDB, для перевода файлов в старый формат была использована программа remediator, установленная на kodomo. Для анализа структур нуклеиновых кислот были использованы программы find_pair и analyze. Программа find_pair определяет спаренные основания и положения спиралей в структуре. В результате работы программ был создан ряд файлов с описанием разных параметров структуры, в файлах 1HDD_old.out и 2DLC_old.out можно найти описание водородных связей, значения всех торсионных углов, ширину малой и большой бороздок и т.д. Упражнение 1. Определение торсионных углов нуклеотидов. В данном упражнении требовалось определить торсионные углы в заданной структуре ДНК и тРНК; с помощью Excel определить среднее значение каждого из торсионных углов (краевые нуклеотиды не рассматривались). Таблица с значениями торсионных углов представлена здесь. На данной таблице представлены средние значения каждого из торсионных углов.
Наиболее "деформированным" нуклеотидом в молекуле ДНК является Т10. Значения торсионных углов α, β, ζ отклоняются от средних значений. В молекуле тРНК - G6. Углы α, β, γ наиболее сильно отклоняются от средних значений. Упражнение 2. Определение структуры водородных связей.
Упражнение 3. Поиск возможных стекинг-взаимодействий. В файле 2DLC.out были найдены данные о величине площади "перекрвывания" двух последовательных пар азотистых оснований. Для пары с наибольшим значением step13 (uP/ga) было получено стандартное изображение стекинг-взаимодействия с помощью программ ex_str и stack2img пакета 3DNA. Результат представлен на рис.6. Рис.6. Изображение стекинг-взаимодействия для набора step13. |
kodomo.fbb.msu.ru
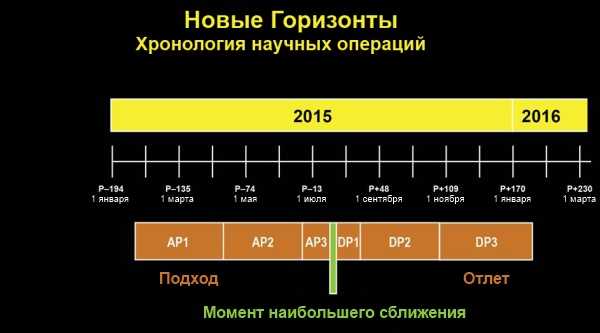
- Космическая миссия новые горизонты
- Корабль корвет фото

- Наша земля круглая или плоская

- Перспективные ракеты носители

- Поиск по фото нейросеть

- Что в ближайшем будущем ждет планету

- Альберт эйнштейн что сделал

- Когда было в последний раз солнечное затмение

- Почему люди невесомы в космосе

- Врожденная нечувствительность к боли
- Холодный ядерный синтез что такое





