Содержание
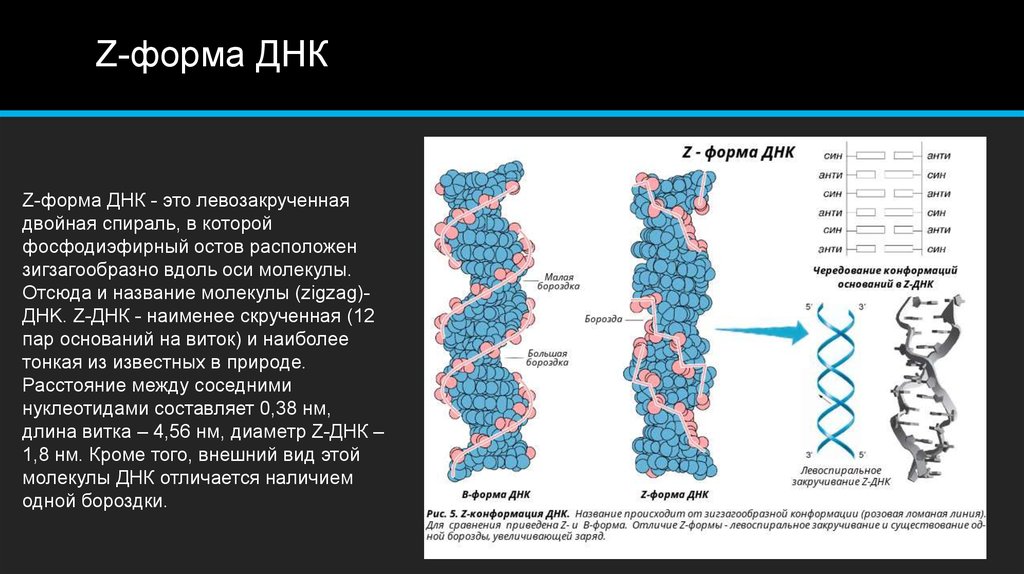
Левозакрученная: загадочная Z-ДНК
Когда-то давно мне пришла в голову идея научно-фантастического романа о жизни на далекой планете, генетический материал которой представлен не правозакрученной, а левозакрученной ДНК. Впрочем, сценарий не такой уж фантастический: левозакрученная ДНК, она же Z-ДНК, есть и в простых земных клетках, хотя функциональное ее предназначение остается малопонятным. Что мы знаем об этой таинственной форме ДНК? Что она делает в живых клетках? Ответам на эти и другие вопросы и посвящен наш обзор.
Слово «ДНК» почти у всех, кто учил в школе биологию, прочно ассоциируется с аккуратной спиральной молекулой, состоящей из двух цепей, закрученных вправо относительно друг друга. Этот образ уже стал элементом массовой культуры: его активно используют в кино и книгах, помещают на логотипы медицинских лабораторий и наклейки на пищевых продуктах, сообщающие, что они «не содержат ГМО» (правда, чаще всего создатели таких логотипов забывают, что ДНК не строго симметрична относительно вертикальной оси, в ней есть большие и малые бороздки). В течение многих лет ученые полагали, что, в отличие от более гибкой РНК, ДНК в клетках представлена исключительно в виде стабильной правозакрученной двойной спирали. Однако исследования последних лет показали, что ДНК не так предсказуема, как кажется. Конечно, по разнообразию возможных пространственных структур до РНК ей далеко, однако в клетке существуют и неканонические формы ДНК: G-квадруплексы, i-мотивы, трехцепочечная ДНК и ряд других элементов, по топологии отличающихся от классической правозакрученной спирали. А еще, как оказалось, двойная спираль ДНК может быть закручена не только вправо, но и влево! Такая левозакрученная ДНК называется Z-ДНК. Несмотря на то, что впервые возможность существования левозакрученной ДНК показали еще в 1979 году [1], Z-ДНК до сих пор окутана ореолом тайны. Так что же мы знаем об этой таинственной форме ДНК?
В течение многих лет ученые полагали, что, в отличие от более гибкой РНК, ДНК в клетках представлена исключительно в виде стабильной правозакрученной двойной спирали. Однако исследования последних лет показали, что ДНК не так предсказуема, как кажется. Конечно, по разнообразию возможных пространственных структур до РНК ей далеко, однако в клетке существуют и неканонические формы ДНК: G-квадруплексы, i-мотивы, трехцепочечная ДНК и ряд других элементов, по топологии отличающихся от классической правозакрученной спирали. А еще, как оказалось, двойная спираль ДНК может быть закручена не только вправо, но и влево! Такая левозакрученная ДНК называется Z-ДНК. Несмотря на то, что впервые возможность существования левозакрученной ДНК показали еще в 1979 году [1], Z-ДНК до сих пор окутана ореолом тайны. Так что же мы знаем об этой таинственной форме ДНК?
Если вы откроете любой учебник по молекулярной биологии, то на первых его страницах наверняка увидите что-то вроде изображенного на рисунке 1:
Рисунок 1. Три основные формы ДНК. а — A-ДНК; б — B-ДНК; в — Z-ДНК.
Три основные формы ДНК. а — A-ДНК; б — B-ДНК; в — Z-ДНК.
Biology:Z-DNA
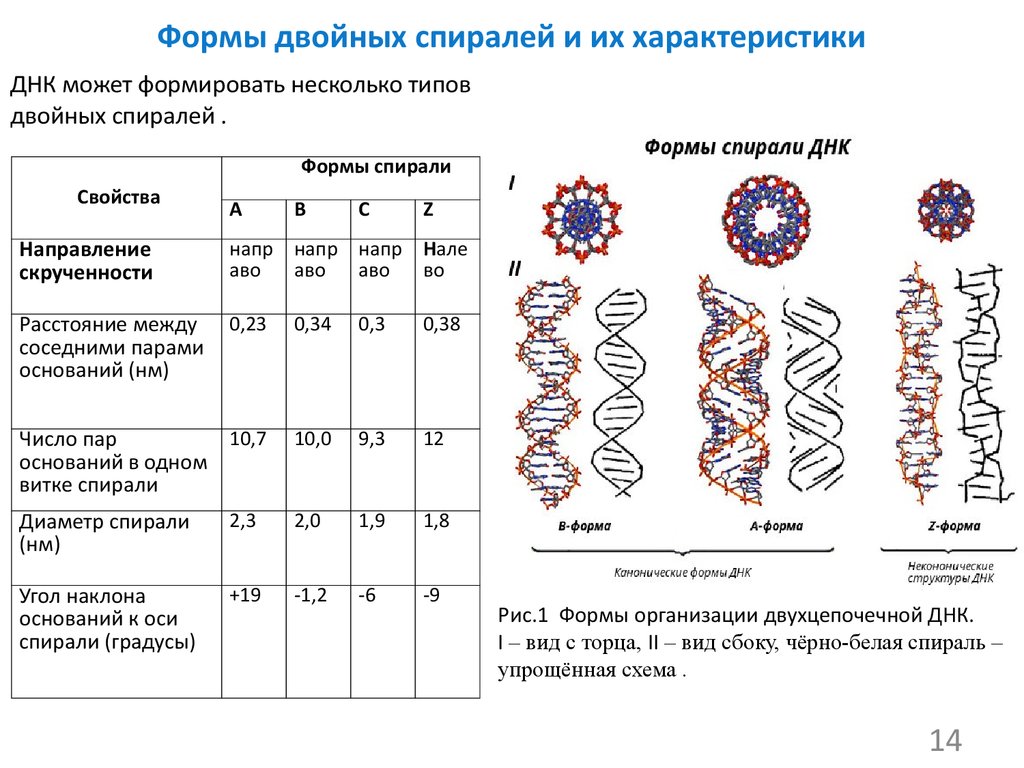
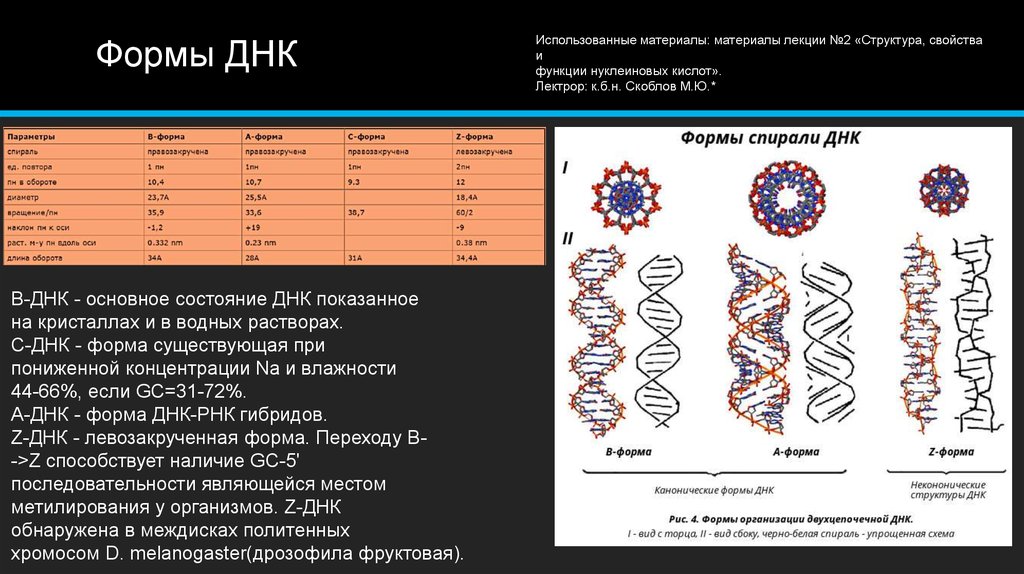
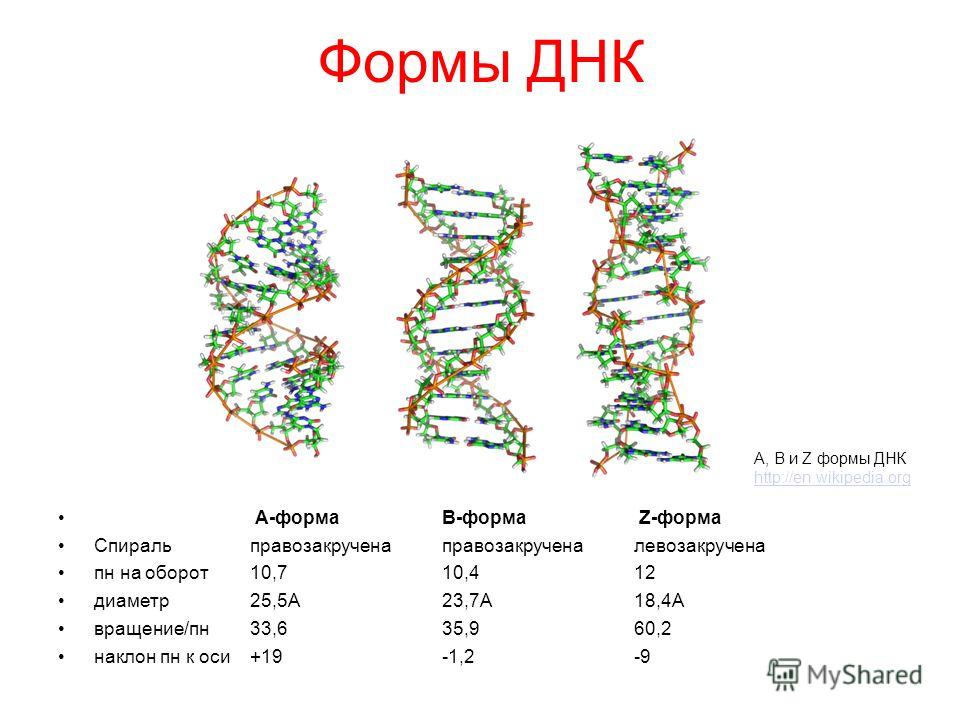
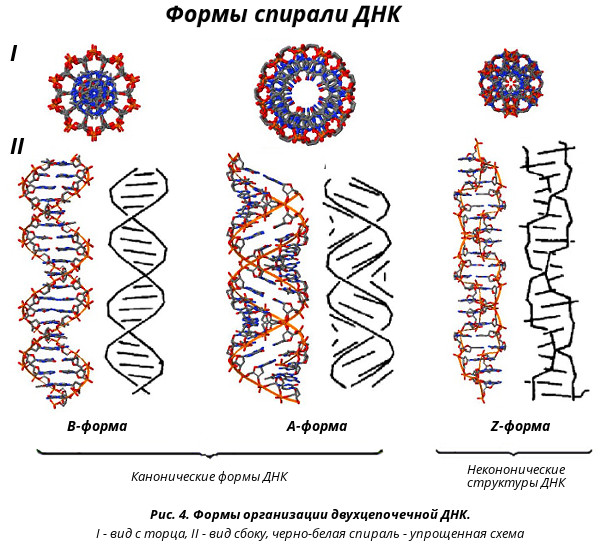
На этой схеме отображены три основные формы ДНК: A-форма, B-форма, которую мы обычно и имеем в виду, говоря о ДНК, и Z-форма — та самая левозакрученная ДНК. Эти формы отличаются друг от друга по множеству геометрических параметров, дотошно измеренных специалистами по кристаллографии и круговому дихроизму (значения этих параметров читатель легко найдет в «Википедии») .
О рентгеноструктурном анализе и других методах структурной биологии вы можете прочитать в нашей статье «12 методов в картинках: структурная биология» [2].
Основные различия между формами таковы: A- и B-ДНК закручены вправо, а Z-ДНК — влево; A-ДНК более «сплющена» по сравнению с B-ДНК, а азотистые основания в ней не перпендикулярны оси спирали, как в B-ДНК. Преобладающая форма, которой и представлен генетический материал живой клетки — B-ДНК, хотя две другие формы, по последним представлениям, также биологически активны. A-форму ДНК получила и описала сама Розалинд Франклин в 1953 году, которая показала, что ДНК переходит в A-форму при дегидратации [3]. В таких условиях геномная ДНК некоторых особо живучих бактерий переходит в A-форму, чтобы хоть как-то сохранить генетическую информацию. Кроме того, в A-форме находится ДНК некоторых вирусов гипертермофильных архей, когда упакована в вирион [4].
A-форму ДНК получила и описала сама Розалинд Франклин в 1953 году, которая показала, что ДНК переходит в A-форму при дегидратации [3]. В таких условиях геномная ДНК некоторых особо живучих бактерий переходит в A-форму, чтобы хоть как-то сохранить генетическую информацию. Кроме того, в A-форме находится ДНК некоторых вирусов гипертермофильных архей, когда упакована в вирион [4].
Z-ДНК была открыта позже A-ДНК и гораздо заметнее отличается от канонической B-формы. Ее впервые описала команда ученых под руководством Роберта Уэлса в 1970 году, когда исследовала полимер, состоящий из повторов инозин—цитозин [5]. Помимо левозакрученности, Z-ДНК отличается также расположением азотистых оснований: в B-форме они находятся в анти-конформации, а в Z-ДНК одно из комплементарных оснований находится в син-конформации, а другое — в анти- (рис. 2).
Рисунок 2. Строение одной пары оснований G:C в составе Z-ДНК. Стрелка указывает на место прохождения оси спирали.
Biology:Z-DNA
За счет этого Z-ДНК выглядит более вытянутой, чем B-ДНК, поскольку расстояние между соседними парами оснований в ней больше, а сахарофосфатный остов по своей конформации напоминает зигзаг. Наконец, стоит отметить, что в Z-ДНК нет большой бороздки [1] (рис. 3).
Рисунок 3. Сравнение B- и Z-ДНК. а — вид сбоку; б — вид сверху.
В отличие от B- и A-форм, которые может принимать ДНК любой нуклеотидной последовательности, Z-ДНК может сформироваться только там, где наблюдается чередование пуринов и пиримидинов. Наибольшая вероятность формирования Z-формы характерна для участков ДНК, состоящих из повторов вида GC, а в области повторов CA и TA существенно ниже. Первоначальные опыты по получению Z-формы ДНК в пробирке проводили в условиях очень высоких концентраций NaCl. Впоследствии оказалось, что искусственно вызвать формирование Z-ДНК можно, добавив в пробирку к B-ДНК комплексные соединения рутения, спермин, спермидин и некоторые другие вещества [1].
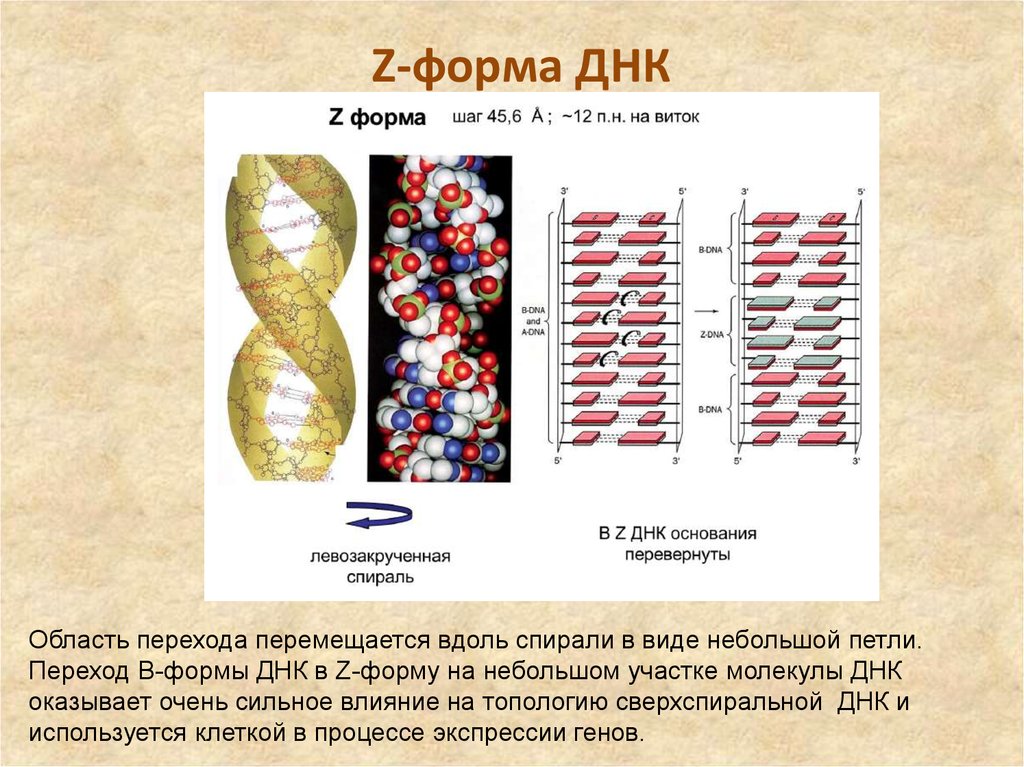
Но может ли Z-ДНК сформироваться в реальных физиологических условиях живой клетки? В 1983 году появилось сообщение, что левозакрученная ДНК присутствует в политенных хромосомах [6]. Кроме того, ранние эксперименты с плазмидной ДНК показали, что формированию Z-ДНК способствует их отрицательная суперскрученность, следовательно, для формирования Z-ДНК не обязательны запредельные концентрации хлорида натрия или другие экзотические соединения: она может сформироваться и в условиях живой клетки. Также выяснили, что образование Z-ДНК стимулируют некоторые белки и модификации оснований. Z-ДНК также может появляться в ходе ремоделирования хроматина при участии комплексов SWI/SNF. Последующие исследования показали, что в тех местах геномной ДНК, где появляется Z-ДНК, на ее границе с обычной B-ДНК образуются неканонические структуры, известные как BZ-соединения (рис. 4). Дело в том, что переход правозакрученной ДНК в левозакрученную — дело энергозатратное и, по большому счету, для клетки невыгодное, и BZ-соединения необходимы для того, чтобы снимать торсионное напряжение в ДНК и тем самым стабилизировать Z-ДНК [1].
Рисунок 4. Структура BZ-соединения. Выпяченные основания на стыке двух конформаций вынесены в отдельное окно и покрашены красным.
Что же касается роли модификаций оснований в формировании Z-ДНК, то в условиях in vitro удалось показать, что метилирование цитозина и гуанина, а также дезаминирование аденина повышают стабильность Z-ДНК. Происходит ли так в реальной клетке — вопрос открытый. Есть наблюдение, что модификации гистонов также влияют на вероятность появления Z-формы ДНК, в частности, ацетилирование гистонов способствует переходу ДНК из B- в Z-форму [1].
Подробнее о модификациях гистонов и других эпигенетических модификациях вы можете прочитать в статьях нашего спецпроекта «Эпигенетика».
Еще один вопрос, не менее сложный и неоднозначный: что полезного Z-ДНК может делать в клетке? Самый очевидный ответ — она может выступать как физический барьер для белков, «ползающих» по ДНК, таких как ДНК- и РНК-полимеразы [1]. В действительности роль Z-ДНК в регуляции транскрипции оказалась не такой однозначной. В одном исследовании 2016 года с помощью иммунопреципитации хроматина с использованием антител к Z-ДНК в геноме человека удалось выявить 191 сайт, склонный к переходу в левозакрученную ДНК [1]. Оказалось, что большинство из этих сайтов находится в промоторах, где Z-ДНК может нивелировать отрицательные супервитки, которые накапливаются позади движущейся по ДНК полимеразы или при освобождении ДНК от нуклеосом. Например, для транскрипции гена, кодирующего колониестимулирующий фактор 1, Z-ДНК строго необходима в области промотора. Кроме того, поскольку Z-ДНК не взаимодействует с нуклеосомами, она остается свободным участком в промоторе, с которым могут связываться транскрипционные факторы. В то же время в случае гена, кодирующего ядрышковый белок нуклеолин, ситуация обратная: появление Z-ДНК в промоторе подавляет экспрессию. Но, пожалуй, наиболее странное наблюдение относительно функций Z-ДНК касается белка E3L вируса коровьей оспы, который летален для мышей.
В действительности роль Z-ДНК в регуляции транскрипции оказалась не такой однозначной. В одном исследовании 2016 года с помощью иммунопреципитации хроматина с использованием антител к Z-ДНК в геноме человека удалось выявить 191 сайт, склонный к переходу в левозакрученную ДНК [1]. Оказалось, что большинство из этих сайтов находится в промоторах, где Z-ДНК может нивелировать отрицательные супервитки, которые накапливаются позади движущейся по ДНК полимеразы или при освобождении ДНК от нуклеосом. Например, для транскрипции гена, кодирующего колониестимулирующий фактор 1, Z-ДНК строго необходима в области промотора. Кроме того, поскольку Z-ДНК не взаимодействует с нуклеосомами, она остается свободным участком в промоторе, с которым могут связываться транскрипционные факторы. В то же время в случае гена, кодирующего ядрышковый белок нуклеолин, ситуация обратная: появление Z-ДНК в промоторе подавляет экспрессию. Но, пожалуй, наиболее странное наблюдение относительно функций Z-ДНК касается белка E3L вируса коровьей оспы, который летален для мышей. Если E3L успешно связывает Z-ДНК, инфекция идет полным ходом и мышь погибает; если связывание заблокировать, то мышь выздоравливает [7].
Если E3L успешно связывает Z-ДНК, инфекция идет полным ходом и мышь погибает; если связывание заблокировать, то мышь выздоравливает [7].
Стоит отметить, что исследования биологических функций Z-ДНК ведутся не очень активно, а некоторые ученые даже считают, что мы переоцениваем биологическое значение левозакрученной ДНК [8]. Работа с Z-ДНК вообще непроста ввиду ее нестабильности в физиологических условиях. В 2019 году китайские ученые предложили, как можно получить Z-ДНК, стабильную в реальных условиях организма. Если взять две замкнутые в кольцо цепи ДНК, комплементарные друг другу, и произвести их гибридизацию в присутствии 0,5 мМ хлорида магния, то получается химерное ZB-кольцо, часть которого будет представлена Z-ДНК, стабильной при физиологических условиях (рис. 5) [9].
Рисунок 5. Схема получения Z-ДНК, стабильной в клеточных условиях. Две кольцевые комплементарные цепи ДНК гибридизуют в присутствии 0,5 мМ хлорида магния, в результате чего получается химерное двухцепочечное кольцо, одна часть которого закручена вправо, а другая — влево.
Еще один механизм формирования Z-ДНК связан с участием белков из группы Z-ДНК-связывающих белков (ZBP). К ним относятся, например, белок E3L вируса коровьей оспы и PKR-подобная протеинкиназа некоторых рыб. У млекопитающих идентифицированы два белка из группы ZBP: аденозиндезаминаза ADAR1 и ДНК-зависимый активатор факторов, регулируемых INF (DAI, также известен как ZBP1). ZBP отличает наличие консервативного домена, связывающего Z-ДНК (его обозначают Zα). Этот домен связывает Z-ДНК чисто за счет конформационного сродства, никаких специфических контактов между основаниями и аминокислотными остатками при этом не образуется. Оба ZBP млекопитающих активируются интерфероном, что указывает на их роль в иммунном ответе против чужеродной ДНК [1]. Как показали недавние исследования, некоторые биологические функции Z-ДНК тесно связаны с ZBP. Например, в 2020 году в Nature появилось сообщение, что распознавание эндогенных Z-нуклеиновых кислот (то есть не только Z-ДНК, но и Z-РНК) белком DAI при помощи Zα-домена запускает некроптоз и воспаление, что может лежать в основе хронических воспалительных заболеваний [10]. Еще одно исследование показало, что митохондриальная ДНК, сильно поврежденная в результате окислительного стресса, может выходить в цитоплазму, где связывается с белком DAI и активирует воспаление. Таким образом, Z-ДНК задействована в регуляции интерферонового ответа [11].
Еще одно исследование показало, что митохондриальная ДНК, сильно поврежденная в результате окислительного стресса, может выходить в цитоплазму, где связывается с белком DAI и активирует воспаление. Таким образом, Z-ДНК задействована в регуляции интерферонового ответа [11].
Как мы уже упоминали, в клетке присутствует не только левозакрученная ДНК, но и левозакрученная РНК (Z-РНК). Ее открыли в 1984 году, показав, что последовательность рибонуклеотидов вида поли-GC переходит в левозакрученную форму в условиях высокой ионной силы и при температуре выше 35 оС [12]. По своим структурным характеристикам Z-РНК близка к Z-ДНК [13]. Существование Z-РНК в живых клетках подтвердили с помощью окрашивания клеток протистов антителами к Z-РНК [14]. Как мы отмечали выше, Zα-домены белков семейства ZBP успешно распознают и Z-РНК тоже, что указывает на ее функциональную связь с белками ADAR1 и DAI [1].
Функциональная связь Z-ДНК с сигнальными путями интерферона свидетельствует, что Z-ДНК иметь отношение к развитию некоторых заболеваний, как минимум, воспалительного характера. Действительно, к настоящему моменту Z-ДНК уже смогли связать с рядом заболеваний человека. Сама по себе Z-ДНК иммунногенна, поэтому она может быть связана с развитием системной красной волчанки, болезни Крона, поликардионеврита и бокового амиотрофического склероза у пациентов, у которых спонтанно начинают образовываться антитела к Z-ДНК. Z-ДНК является фактором структурной нестабильности генома, поэтому она может лежать в основе онкологических заболеваний, особенно если геномные перестройки затрагивают такие гены, как BCL2, C-MYC и SCL. Z-ДНК может формироваться в области повторов CGG, связанных с синдромом ломкой X-хромосомы, и повторов GAC, которые связывают со скелетной дисплазией. Наконец, показано, что у пациентов с болезнью Альцгеймера в нейронах гиппокампа выявляется аномальное количество Z-ДНК, что связывает ее и с нейродегенеративными заболеваниями [1].
Действительно, к настоящему моменту Z-ДНК уже смогли связать с рядом заболеваний человека. Сама по себе Z-ДНК иммунногенна, поэтому она может быть связана с развитием системной красной волчанки, болезни Крона, поликардионеврита и бокового амиотрофического склероза у пациентов, у которых спонтанно начинают образовываться антитела к Z-ДНК. Z-ДНК является фактором структурной нестабильности генома, поэтому она может лежать в основе онкологических заболеваний, особенно если геномные перестройки затрагивают такие гены, как BCL2, C-MYC и SCL. Z-ДНК может формироваться в области повторов CGG, связанных с синдромом ломкой X-хромосомы, и повторов GAC, которые связывают со скелетной дисплазией. Наконец, показано, что у пациентов с болезнью Альцгеймера в нейронах гиппокампа выявляется аномальное количество Z-ДНК, что связывает ее и с нейродегенеративными заболеваниями [1].
К сожалению, наши знания о биологии Z-ДНК и тем более Z-РНК весьма обрывочны и неполны. Остается надеяться, что новые исследования позволят нам узнать больше об этих таинственных молекулах.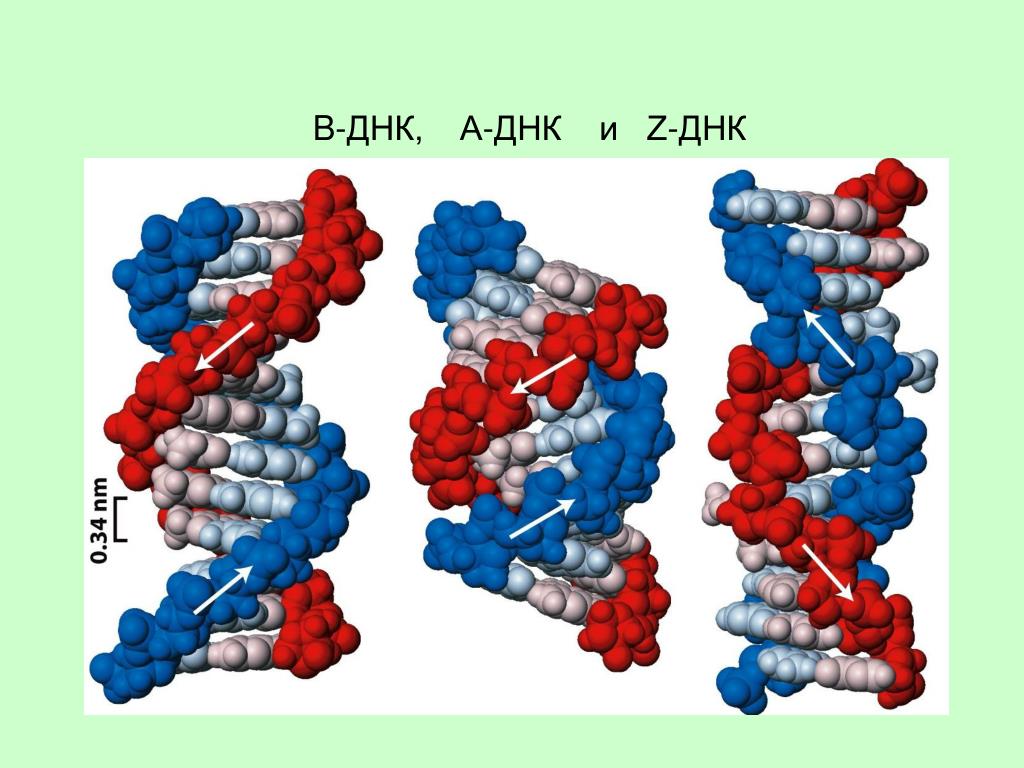 А может быть, когда-нибудь на полках магазинов появится научно-фантастический роман, посвященный жизни, основанной на левозакрученной ДНК…
А может быть, когда-нибудь на полках магазинов появится научно-фантастический роман, посвященный жизни, основанной на левозакрученной ДНК…
- Subramaniyam Ravichandran, Vinod Kumar Subramani, Kyeong Kyu Kim. (2019). Z-DNA in the genome: from structure to disease. Biophys Rev. 11, 383-387;
- 12 методов в картинках: структурная биология;
- R. E. Franklin, R. G. Gosling. (1953). The structure of sodium thymonucleate fibres. I. The influence of water content. Acta Crystallogr. 6, 673-677;
- F. DiMaio, X. Yu, E. Rensen, M. Krupovic, D. Prangishvili, E. H. Egelman. (2015). A virus that infects a hyperthermophile encapsidates A-form DNA. Science. 348, 914-917;
- Y. MITSUI, R. LANGRIDGE, BARBARA E. SHORTLE, CHARLES R. CANTOR, ROBERT C. GRANT, et. al.. (1970). Physical and Enzymatic Studies on Poly d(I–C).Poly d(I–C), an Unusual Double-helical DNA.
 Nature. 228, 1166-1169;
Nature. 228, 1166-1169; - Michel Robert-Nicoud, Donna J. Arndt-Jovin, David A. Zarling, Thomas M. Jovin. (1983). LEFT-HANDED Z DNA IN POLYTENE CHROMOSOMES. Mobility and recognition in cell biology. 281-290;
- Sung Chul Ha, Ky Lowenhaupt, Alexander Rich, Yang-Gyun Kim, Kyeong Kyu Kim. (2005). Crystal structure of a junction between B-DNA and Z-DNA reveals two extruded bases. Nature. 437, 1183-1186;
- Alan Herbert. (2019). Z-DNA and Z-RNA in human disease. Commun Biol. 2;
- Yaping Zhang, Yixiao Cui, Ran An, Xingguo Liang, Qi Li, et. al.. (2019). Topologically Constrained Formation of Stable Z-DNA from Normal Sequence under Physiological Conditions. J. Am. Chem. Soc.. 141, 7758-7764;
- Huipeng Jiao, Laurens Wachsmuth, Snehlata Kumari, Robin Schwarzer, Juan Lin, et. al.. (2020). Z-nucleic-acid sensing triggers ZBP1-dependent necroptosis and inflammation. Nature. 580, 391-395;
- Bartosz Szczesny, Michela Marcatti, Akbar Ahmad, Mauro Montalbano, Attila Brunyánszki, et. al.. (2018). Mitochondrial DNA damage and subsequent activation of Z-DNA binding protein 1 links oxidative stress to inflammation in epithelial cells. Sci Rep. 8;
- Kathleen Hall, Phillip Cruz, Ignacio Tinoco, Thomas M. Jovin, Johan H. van de Sande. (1984). ‘Z-RNA’—a left-handed RNA double helix. Nature. 311, 584-586;
- M. Popenda. (2004). High salt solution structure of a left-handed RNA double helix. Nucleic Acids Research. 32, 4044-4054;
- D. A. Zarling, C. J. Calhoun, C. C. Hardin, A. H. Zarling. (1987). Cytoplasmic Z-RNA.. Proceedings of the National Academy of Sciences. 84, 6117-6121;
- Doyoun Kim, Sanjith Reddy, Dong Young Kim, Alexander Rich, Sangho Lee, et. al.. (2009). Base extrusion is found at helical junctions between right- and left-handed forms of DNA and RNA. Nucleic Acids Research. 37, 4353-4359.
 Nature. 228, 1166-1169;
Nature. 228, 1166-1169;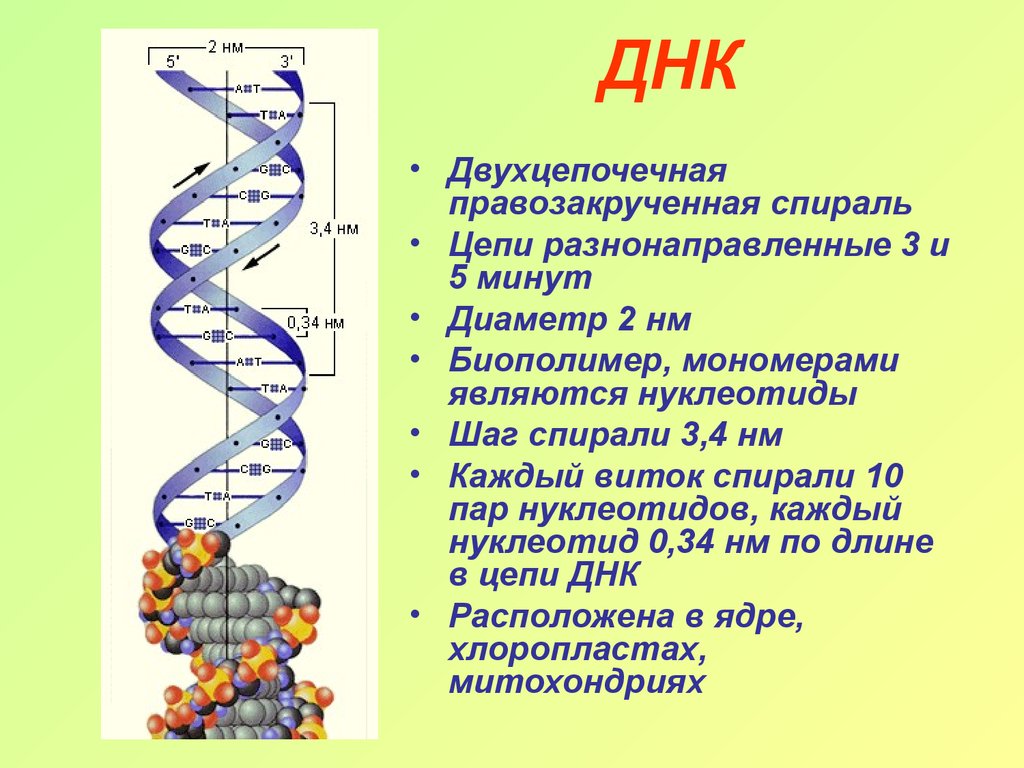 (2020). Z-nucleic-acid sensing triggers ZBP1-dependent necroptosis and inflammation. Nature. 580, 391-395;
(2020). Z-nucleic-acid sensing triggers ZBP1-dependent necroptosis and inflammation. Nature. 580, 391-395;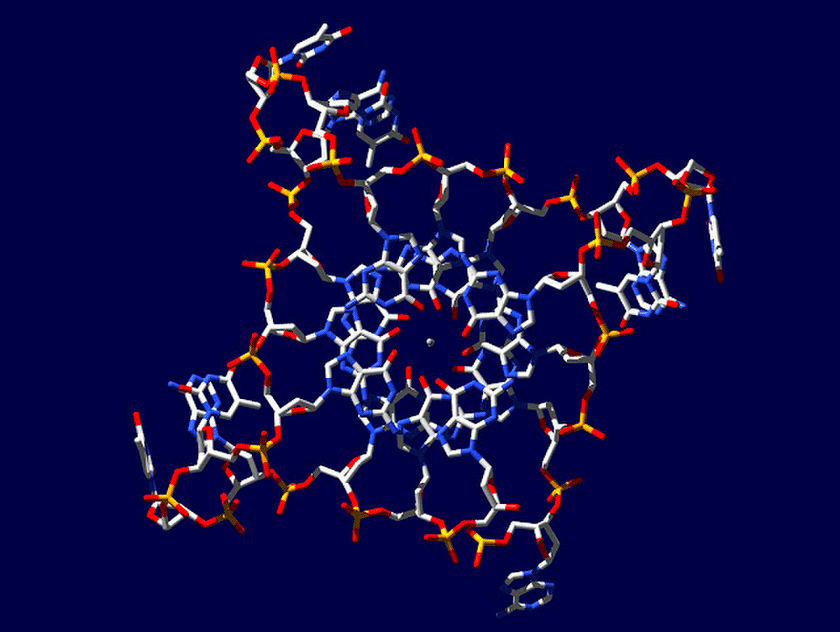 al.. (2009). Base extrusion is found at helical junctions between right- and left-handed forms of DNA and RNA. Nucleic Acids Research. 37, 4353-4359.
al.. (2009). Base extrusion is found at helical junctions between right- and left-handed forms of DNA and RNA. Nucleic Acids Research. 37, 4353-4359.ДНК • Джеймс Трефил, энциклопедия «Двести законов мироздания»
Молекула ДНК имеет форму двойной спирали, и ее воспроизведение основано на том, что каждая цепь двойной спирали служит матрицей для сборки новых молекул.
Сегодня мы знаем, что молекула ДНК является носителем кода, который управляет химизмом всего живого (см. Центральная догма молекулярной биологии), а двойная спираль молекулы ДНК стала одним из самых известных научных символов. Открытие ДНК, как и практически все великие открытия, не было результатом работы одинокого гения, а увенчало собой длинную цепь экспериментальных работ. Так, эксперимент Херши—Чейз продемонстрировал, что носителем генетической информации в клетках является именно ДНК, а не белки. Еще в 1920-е годы американский биохимик родом из России Фибус Левин (Phoebus Levene, 1869–1940) установил, что основные кирпичики, из которых построена ДНК, — это пятиатомный сахар дезоксирибоза (она обозначена буквой Д в слове ДНК), фосфатная группа и четыре азотистых основания — тимин, гуанин, цитозин и аденин (их обычно обозначают буквами Т, Г, Ц и А).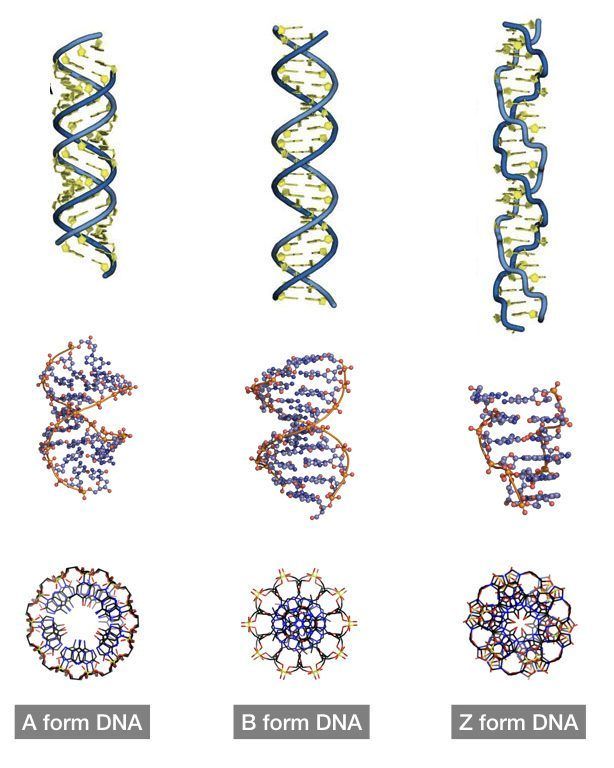 В конце 1940-х годов американский биохимик австрийского происхождения Эрвин Чаргафф (Erwin Chargaff, р. 1905) выяснил, что во всех ДНК содержится равное количество оснований Т и А и, аналогично, равное количество оснований Г и Ц. Однако относительное содержание Т/А и Г/Ц в молекуле ДНК специфично для каждого вида.
В конце 1940-х годов американский биохимик австрийского происхождения Эрвин Чаргафф (Erwin Chargaff, р. 1905) выяснил, что во всех ДНК содержится равное количество оснований Т и А и, аналогично, равное количество оснований Г и Ц. Однако относительное содержание Т/А и Г/Ц в молекуле ДНК специфично для каждого вида.
В начале 1950-х годов стали известны два новых факта, пролившие свет на природу ДНК: американский химик Лайнус Полинг (Linus Pauling, 1901–94) показал, что в длинных молекулах, например белках, могут образовываться связи, закручивающие молекулу в спираль, а в лондонской лаборатории Морис Уилкинс и Розалинда Франклин получили данные рентгеноструктурного анализа (основанные на усовершенствованном применении закона Брэгга), позволившие предположить, что ДНК имеет спиральную структуру.
Как раз в это время молодой американский биохимик Джеймс Уотсон отправился на год в Кембриджский университет для работы с молодым английским физиком-теоретиком Фрэнсисом Криком. («Обо мне тогда практически никто не знал, — вспоминал впоследствии Крик, — а идеи Уотсона считали. .. слишком заумными».) Экспериментируя с металлическими моделями, Крик и Уотсон пытались объединить различные компоненты молекулы в трехмерную модель ДНК.
.. слишком заумными».) Экспериментируя с металлическими моделями, Крик и Уотсон пытались объединить различные компоненты молекулы в трехмерную модель ДНК.
Чтобы лучше представить себе полученные ими результаты, вообразите длинную лестницу. Вертикальные стойки этой лестницы состоят из молекул сахара, кислорода и фосфора. Важную функциональную информацию в молекуле несут ступеньки лестницы. Они состоят из двух молекул, каждая из которых крепится к одной из вертикальных стоек. Эти молекулы — четыре азотистых основания — представляют собой одиночные или двойные кольца, содержащие атомы углерода, азота и кислорода и способные образовывать две или три водородные связи (см. Химические связи) с другими основаниями. Форма этих молекул позволяет им образовывать связи — законченные ступеньки — лишь определенного типа: между А и Т и между Г и Ц. Другие связи возникнуть не могут. Следовательно, каждая ступенька представлена либо А—Т либо Г—Ц. Теперь вообразите, что вы берете собранную таким образом лестницу за два конца и скручиваете — вы получите знакомую двойную спираль ДНК.
Считывая ступеньки по одной цепи молекулы ДНК, вы получите последовательность оснований. Представьте, что это сообщение, написанное с помощью алфавита всего из четырех букв. Именно это сообщение определяет химические превращения, происходящие в клетке, и, следовательно, характеристики живого организма, частью которого является эта клетка. На другой цепи спирали никакой новой информации не содержится, ведь если вам известно основание, которое находится на одной цепи, вы знаете и то, какой должна быть вторая половина ступеньки. В некотором смысле две цепи двойной спирали относятся друг другу так же, как фотография и негатив.
Открыв двуспиральную структуру ДНК, Уотсон и Крик поняли и тот простой способ, которым осуществляется воспроизведение молекулы ДНК — как и должно происходить при делении клетки. По их собственным словам, «от нашего внимания не ускользнул тот факт, что постулированная нами специфичная парность азотистых оснований непосредственно указывает на возможный механизм копирования генетического материала».
Такой «возможный механизм копирования» определен структурой ДНК. Когда клетка приступает к делению и необходима дополнительная ДНК для дочерних клеток, ферменты (см. Катализаторы и ферменты) начинают «расстегивать» лестницу ДНК, как застежку-«молнию», обнажая индивидуальные основания. Другие ферменты присоединяют соответствующие основания, находящиеся в окружающей жидкой среде, к парным «обнажившимся» основаниям — А к Т, Г к Ц и т. д. В результате на каждой из двух разошедшихся цепей ДНК достраивается соответствующая ей цепь из компонентов окружающей среды, и исходная молекула дает начало двум двойным спиралям.
Точно так же, как каждое великое открытие основано на работе предшественников, оно дает начало новым плодотворным исследованиям, поскольку ученые используют полученную информацию для движения вперед. Можно сказать, что открытие двойной спирали дало толчок последующему полувековому развитию молекулярной биологии, завершившемуся успешным осуществлением проекта «Геном человека».
2.5: B-форма, A-форма и Z-форма ДНК
- Последнее обновление
- Сохранить как PDF
- Идентификатор страницы
- 307
- Росс Хардисон
- Университет штата Пенсильвания
Три основные формы ДНК являются двухцепочечными и связаны взаимодействиями между комплементарными парами оснований. Это термины A-форма, B-форма и Z-форма ДНК.
ДНК B-формы
Информация о базовом составе ДНК, знание структуры динуклеотидов и понимание того, что рентгеновская кристаллография предполагает наличие спиральной периодичности, были объединены Уотсоном и Криком в 1953 г. в предложенной ими модели двойного спиральная структура ДНК. Они предложили две нити ДНК — каждая в виде правосторонней спирали — закручены вокруг одной и той же оси. Две нити удерживаются вместе Н-связями между основаниями (в антиконформации), как показано на рисунке \(\PageIndex{1}\).
Большая бороздка Большая бороздка
Малая бороздка Малая бороздка
Рисунок \(\PageIndex{1}\): (слева) Пара оснований A:T и (справа) пара оснований G:C
Основания соответствуют модели двойной спирали, если пиримидин на одной нити всегда сочетается с пурином на другой. Согласно правилам Чаргаффа , две нити будут соединять А с Т и G с С. Это связывает кетооснование с аминооснованием, пурин с пиримидином. Между A и T могут образоваться две Н-связи, а между G и C могут образоваться три Н-связи. Эта третья Н-связь в паре оснований G:C находится между дополнительной экзоциклической аминогруппой на G и кетогруппой C2 на C. Кетогруппа пиримидина С2 не участвует в образовании водородных связей в паре оснований А:Т.
Это комплементарные пары оснований. Схема спаривания оснований сразу предлагает способ репликации и копирования генетической информации.
Рисунок \(\PageIndex{2}\): Антипараллельные (а), плектонемически закрученные (б, в, г) нити ДНК. Стрелки на а указывают от 3’ до 5’, но они иллюстрируют антипараллельный характер дуплекса. Две нити дуплекса антипараллельны и плектонемически закручены. Нуклеотиды, расположенные в ориентации от 5′ до 3′ на одной цепи, выравниваются с комплементарными нуклеотидами в ориентации от 3′ до 5′ противоположной нити.
Две нити расположены не просто рядом друг с другом, что можно было бы назвать паранемическим соединением (рис. \(\PageIndex{3}\)). (С этим мы столкнемся во время рекомбинации в Главе 8.) Скорее две нити закручены вокруг одной и той же оси спирали и переплетены друг с другом (что называется плектонемической спиралью ). Одним из следствий этого переплетения является то, что две нити не могут быть разделены без вращения ДНК, один оборот ДНК на каждое «раскручивание» двух нитей.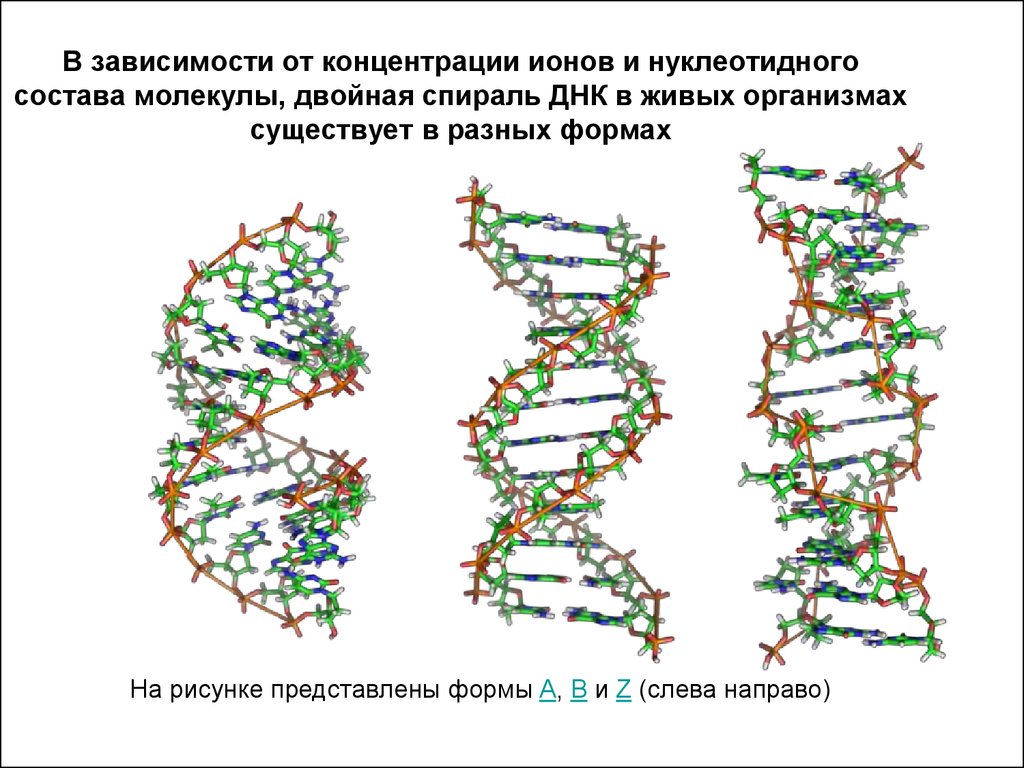
Рисунок \(\PageIndex{3}\): Две нити дуплекса ДНК закручены друг вокруг друга в виде плектонемического витка (слева), а не паранемического дуплекса (справа).
Размеры В-формы (наиболее распространенной) ДНК
- 0,34 нм между п.н., 3,4 нм на виток, около 10 п.н. на виток
- 1,9 нм (около 2,0 нм или 20 Ангстрем) в диаметре
Большая и малая бороздки
Большая бороздка шире, чем малая бороздка в ДНК (рис. \(\PageIndex{2d}\)), и в большой бороздке взаимодействует множество специфичных для последовательности белков. Группы N7 и C6 пуринов и группы C4 и C5 пиримидинов обращены в большую бороздку, поэтому они могут устанавливать специфические контакты с аминокислотами в ДНК-связывающих белках. Таким образом, определенные аминокислоты служат донорами и акцепторами Н-связей для образования Н-связей с определенными нуклеотидами в ДНК. Доноры и акцепторы Н-связи также находятся в малой бороздке, и действительно, некоторые белки связываются именно в малой бороздке. Пары оснований складываются с некоторым вращением между ними.
Пары оснований складываются с некоторым вращением между ними.
Нуклеиновые кислоты A-формы и Z-ДНК
Описаны три различные формы дуплексных нуклеиновых кислот. Наиболее распространенной формой, присутствующей в большинстве ДНК при нейтральном pH и физиологических концентрациях солей, является B-форма. Это классическая правосторонняя двойная спиральная структура, которую мы обсуждали. Более толстый правый дуплекс с более коротким расстоянием между парами оснований был описан для дуплексов РНК-ДНК и дуплексов РНК-РНК. Это называется А-форма нуклеиновой кислоты.
Третья форма дуплексной ДНК имеет поразительно отличающуюся левостороннюю спиральную структуру. Эта Z-ДНК образована чередующимися участками пуринов и пиримидинов, т.е. GCGCGC, особенно в отрицательно суперскрученной ДНК. Небольшое количество ДНК в клетке существует в Z-форме. Было заманчиво предположить, что эта другая структура каким-то образом участвует в регуляции некоторых клеточных функций, таких как транскрипция или регуляция, но убедительных доказательств за или против этого предположения пока нет.
Различия между А-формой и В-формой нуклеиновой кислоты
Основное различие между А-формой и В-формой нуклеиновой кислоты заключается в конформации дезоксирибозного сахарного кольца. Он находится в эндоконформации C2′ для B-формы, тогда как он находится в эндоконформации C3′ в A-форме. Как показано на рисунке \(\PageIndex{4}\), если вы рассматриваете плоскость, определяемую атомами C4′-O-C1′ дезоксирибозы, в эндоконформации C2′ атом C2′ находится над плоскостью, тогда как атом C3′ находится над плоскостью в эндоконформации C3′. Последняя конформация сближает 5′- и 3′-гидроксилы (оба этерифицированы фосфатами, связывающимися со следующими нуклеотидами) ближе друг к другу, чем в С2′-эндоконформации (рис. 2.16). Таким образом, расстояние между соседними нуклеотидами уменьшается примерно на 1 ангстрем в А-форме по сравнению с В-формой нуклеиновой кислоты (рис. \(\PageIndex{4}\)).
Рисунок \(\PageIndex{4}\): Син- и анти-конформации основания по отношению к сахару в нуклеотидах.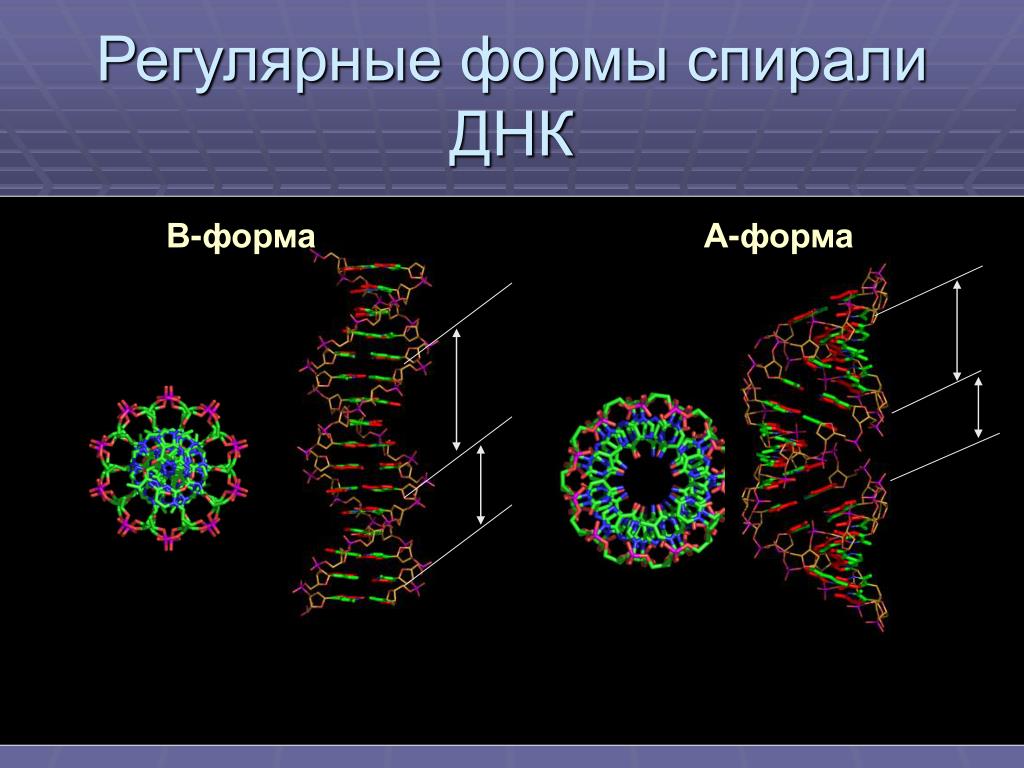
Второе важное различие между нуклеиновой кислотой A-формы и B-формы заключается в расположении пар оснований в дуплексе. В форме B пары оснований расположены почти по центру оси спирали (рис. \(\PageIndex{4}\)), а в форме A они смещены от центральной оси и ближе к большой бороздке. В результате получается лентообразная спираль с более открытой цилиндрической сердцевиной А-формы.
Z-форма ДНК
Z-ДНК представляет собой радикально отличающуюся дуплексную структуру с двумя нитями, закрученными в левые спирали, и выраженным зигзагообразным (отсюда и название) рисунком в фосфодиэфирном остове. Как упоминалось ранее, Z-ДНК может образовываться, когда ДНК находится в чередующейся пурин-пиримидиновой последовательности, такой как GCGCGC, и действительно, нуклеотиды G и C находятся в разных конформациях, что приводит к зигзагообразной структуре. Большая разница в нуклеотиде G. Он имеет сахар в эндоконформации C3′ (как нуклеиновая кислота A-формы и в отличие от ДНК B-формы), а основание гуанина находится в синконформации.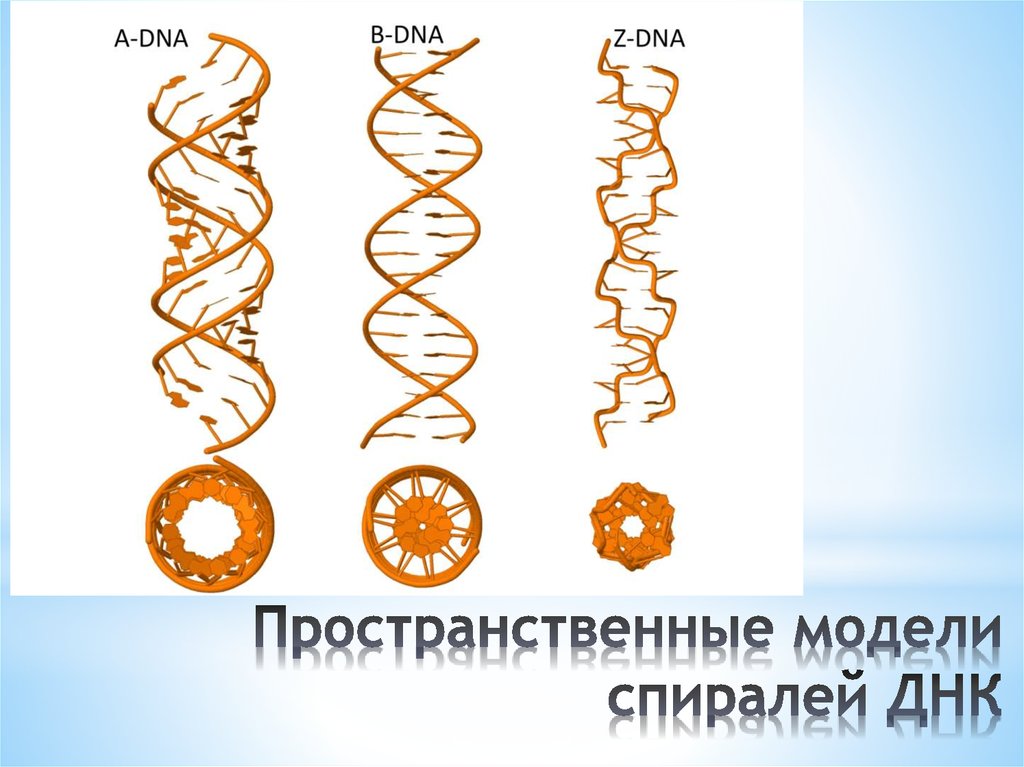 Это помещает гуанин обратно поверх сахарного кольца, в отличие от обычной антиконформации, наблюдаемой в нуклеиновых кислотах A- и B-форм. Обратите внимание, что наличие основания в антиконформации помещает его в положение, в котором оно может легко образовывать Н-связи с комплементарным основанием на противоположной цепи. Дуплекс в Z-ДНК должен приспосабливаться к искажению этого нуклеотида G в синконформации. Цитозин в соседнем нуклеотиде Z-ДНК находится в «нормальном» С2′-эндо, антиконформации.
Это помещает гуанин обратно поверх сахарного кольца, в отличие от обычной антиконформации, наблюдаемой в нуклеиновых кислотах A- и B-форм. Обратите внимание, что наличие основания в антиконформации помещает его в положение, в котором оно может легко образовывать Н-связи с комплементарным основанием на противоположной цепи. Дуплекс в Z-ДНК должен приспосабливаться к искажению этого нуклеотида G в синконформации. Цитозин в соседнем нуклеотиде Z-ДНК находится в «нормальном» С2′-эндо, антиконформации.
Рисунок \(\PageIndex{5}\): B-форма (слева), A-форма (в центре) и Z-ДНК (справа). (CC BY-SA 4.0; Mauroesguerroto)
Даже классическая B-ДНК не является полностью однородной по своей структуре. Рентгеноструктурный анализ кристаллов дуплексных олигонуклеотидов показывает, что данная последовательность будет иметь отличительную структуру. Эти вариации в B-ДНК могут различаться поворотом пропеллера (между основаниями в паре) для оптимизации укладки оснований или тремя способами, которыми 2 последовательные пары оснований могут перемещаться относительно друг друга: скручивание, перекатывание или скольжение.
| Форма B | Форма А | Z-форма | |
|---|---|---|---|
| смысл спирали | Правша | Правша | Левша |
| пар оснований на оборот | 10 | 11 | 12 |
| вертикальный подъем на бп | 3,4 Å | 2,56 Å | 19 Å |
| оборот на бит | +36° | +33° | -30° |
| диаметр спирали | 19 Å | 19 Å | 19 Å |
Эта страница под названием 2.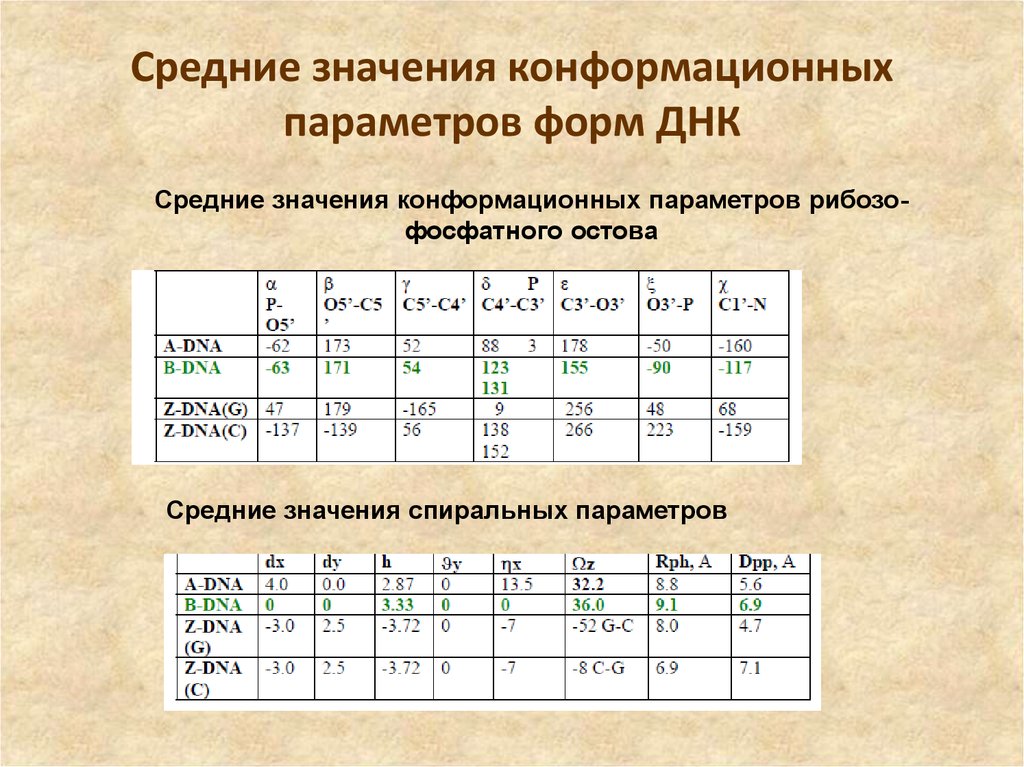 5: B-форма, A-форма и Z-форма ДНК распространяется по незаявленной лицензии и была создана, изменена и/или курирована Россом Хардисоном.
5: B-форма, A-форма и Z-форма ДНК распространяется по незаявленной лицензии и была создана, изменена и/или курирована Россом Хардисоном.
- Наверх
- Была ли эта статья полезной?
- Тип изделия
- Раздел или страница
- Автор
- Росс Хардисон
- Показать оглавление
- нет
- Теги
- Правила Чаргаффа
- ДНК (форма А)
- ДНК (форма B)
- ДНК (форма Z)
- плектонемическая катушка
Различные формы ДНК – форма A, форма B, форма Z
Сагар Арьял
- Правосторонняя двойная спираль Watson – Crick Model для B-формы ДНК является наиболее известной структурой ДНК.
- В дополнение к этой классической структуре наблюдалось несколько других форм ДНК.
- Таким образом, спиральная структура ДНК изменчива и зависит как от последовательности, так и от окружающей среды.
Содержание
Почему существуют разные формы ДНК?
- ДНК просто не хватает места, чтобы растянуть ее в идеальную линейную конформацию B-ДНК. Почти во всех клетках, от простых бактерий до сложных эукариот, ДНК должна быть сжата более чем в тысячу раз, чтобы даже поместиться внутри клетки или ядра.
- Уточненное разрешение структуры ДНК, основанное на рентгеновской кристаллографии коротких синтетических фрагментов ДНК, показало, что спиральная структура ДНК значительно различается в зависимости от последовательности. Например, участок ДНК размером 200 п.н. может работать так, как если бы он имел длину более 1000 п.н. на акриламидном геле, если он имеет правильную последовательность. Двойная спираль не является такой же однородной структурой.
 Например, участок ДНК размером 200 п.н. может работать так, как если бы он имел длину более 1000 п.н. на акриламидном геле, если он имеет правильную последовательность. Двойная спираль не является такой же однородной структурой.
Например, участок ДНК размером 200 п.н. может работать так, как если бы он имел длину более 1000 п.н. на акриламидном геле, если он имеет правильную последовательность. Двойная спираль не является такой же однородной структурой.Различные формы ДНК
ДНК B-формы
- B-ДНК представляет собой форму двойной спирали Уотсона-Крика, с которой знакомо большинство людей.
- Они предложили две нити ДНК — каждая в виде правосторонней спирали — закручены вокруг одной оси. Две нити удерживаются вместе Н-связями между основаниями (в антиконформации).
- Две нити дуплекса антипараллельны и закручены плектонемически. Нуклеотиды, расположенные с ориентацией от 5′ к 3′ на одной цепи, выравниваются с комплементарными нуклеотидами с ориентацией от 3′ к 5′ противоположной нити.
- Основания соответствуют модели двойной спирали, если пиримидин на одной цепи всегда сочетается с пурином на другой. Согласно правилам Чаргаффа, две цепи будут соединять А с Т и G с Ц. Это связывает кетооснование с аминооснованием, пурин с пиримидином. Две Н-связи могут образовываться между А и Т, а три — между G и С.
- Это комплементарные пары оснований. Схема спаривания оснований сразу предлагает способ репликации и копирования генетической информации.
- 34 нм между п.н., 3,4 нм на виток, около 10 п.н. на виток
- 9 нм (около 2,0 нм или 20 Ангстрем) в диаметре.
- 34 o шаг спирали; -6 o наклон пары оснований; 36 o Угол поворота
 Это связывает кетооснование с аминооснованием, пурин с пиримидином. Две Н-связи могут образовываться между А и Т, а три — между G и С.
Это связывает кетооснование с аминооснованием, пурин с пиримидином. Две Н-связи могут образовываться между А и Т, а три — между G и С.ДНК А-формы
- Основное различие между нуклеиновой кислотой А-формы и В-формы заключается в подтверждении присутствия дезоксирибозного сахарного кольца. Он находится в эндоконформации C2′ для B-формы, тогда как он находится в эндоконформации C3′ в A-форме.
- Второе важное различие между нуклеиновой кислотой формы A и формы B заключается в размещении пар оснований в дуплексе. В форме B пары оснований почти центрированы относительно оси спирали, но в форме A они смещены от центральной оси и ближе к большой бороздке. В результате получается лентообразная спираль с более открытой цилиндрической сердцевиной А-формы.
- Правосторонняя спираль
- 11 бп за ход; осевой подъем 0,26 нм; 28 o шаг спирали; 20 o наклон пары оснований
- 33 o Угол поворота; Диаметр спирали 2,3 нм
 В результате получается лентообразная спираль с более открытой цилиндрической сердцевиной А-формы.
В результате получается лентообразная спираль с более открытой цилиндрической сердцевиной А-формы.Z-форма ДНК
- Z-ДНК представляет собой радикально отличающуюся дуплексную структуру с двумя нитями, закрученными в левые спирали, и выраженным зигзагообразным рисунком (отсюда и название) в фосфодиэфирном остове.
- Z-ДНК может образовываться, когда ДНК находится в чередующейся пурин-пиримидиновой последовательности, такой как GCGCGC, и действительно, нуклеотиды G и C находятся в разных конформациях, что приводит к зигзагообразному рисунку.
- Большая разница в нуклеотиде G.
- Он имеет сахар в эндоконформации C3′ (как нуклеиновая кислота в форме A и в отличие от ДНК в форме B), а основание гуанина находится в синконформации.
- Это помещает гуанин обратно поверх сахарного кольца, в отличие от обычной антиконформации, наблюдаемой в нуклеиновых кислотах A- и B-форм. Обратите внимание, что наличие основания в антиконформации помещает его в положение, в котором оно может легко образовывать Н-связи с комплементарным основанием на противоположной цепи.
- Дуплекс в Z-ДНК должен компенсировать искажение этого нуклеотида G в синконформации. Цитозин в соседнем нуклеотиде Z-ДНК находится в «нормальном» С2′-эндо, антиконформации.
- Обнаружен Ричем, Нордхеймом и Вангом в 1984 году.
- Имеет антипараллельные нити, как В-ДНК.
- Длинная и тонкая по сравнению с В-ДНК.
- 12 бп за ход; подъем по оси 0,45 нм; 45 o шаг спирали; 7 o наклон пары оснований
- -30 o Угол поворота; Диаметр спирали 1,8 нм
Условия, благоприятствующие A-форме, B-форме и Z-форме ДНК
- Будет ли последовательность ДНК находиться в A-, B- или Z-форме ДНК, зависит как минимум от трех условий.
- Во-первых, это ионная или гидратационная среда, которая может облегчить преобразование между различными спиральными формами.
- Для A-ДНК благоприятствует низкая гидратация, тогда как для Z-ДНК может благоприятствовать высокое содержание соли.
- Вторым условием является последовательность ДНК: A-ДНК благоприятствуют определенным участкам пуринов (или пиримидинов), тогда как Z-ДНК легче всего может быть образована путем чередования пурин-пиримидиновых стадий.
- Третьим условием является наличие белков, которые могут связываться с ДНК в одной спиральной конформации и заставлять ДНК принимать другую конформацию, таких как белки, которые связываются с B-ДНК и могут привести ее к A- или Z-формам.
- В живых клетках большая часть ДНК находится в смеси A- и B-конформаций ДНК с несколькими небольшими участками, способными образовывать Z-ДНК.
Другие редкие формы ДНК. Д-ДНК E-ДНК Ссылки