Содержание
Рудименты и атавизмы. — Датированный ежедневник. Записная книжка. — ЖЖ
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


 Таким образом, специфическая сложность крыльев страуса неадекватна простоте задач, для которых эти крылья используются, и именно поэтому эти крылья называют рудиментами. В числе других нелетающих птиц с рудиментарными крыльями — галапагосские бакланы (Phalacrocorax harrisi), киви и какапо. Для сравнения, крыло пингвина имеет большое значение, действуя в качестве плавника, а значит — не может считаться рудиментом.»
Таким образом, специфическая сложность крыльев страуса неадекватна простоте задач, для которых эти крылья используются, и именно поэтому эти крылья называют рудиментами. В числе других нелетающих птиц с рудиментарными крыльями — галапагосские бакланы (Phalacrocorax harrisi), киви и какапо. Для сравнения, крыло пингвина имеет большое значение, действуя в качестве плавника, а значит — не может считаться рудиментом.» Она прикрепляется к копчику, но, поскольку копчик у человека практически не может двигаться, эта мышца для человека бесполезна.
Она прикрепляется к копчику, но, поскольку копчик у человека практически не может двигаться, эта мышца для человека бесполезна.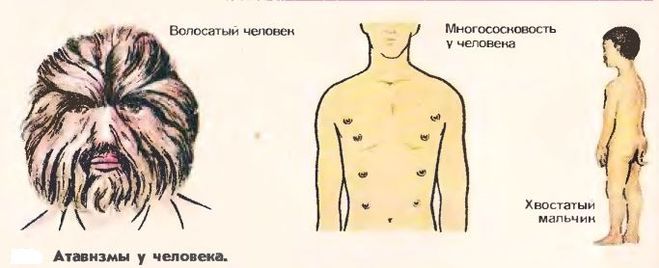
 Многие другие виды насекомых, недавно утратившие способность к полету, такие как водяной скорпион Nepa cinerea, имеют внешне вполне нормальные крылья, которыми никогда не пользуются.
Многие другие виды насекомых, недавно утратившие способность к полету, такие как водяной скорпион Nepa cinerea, имеют внешне вполне нормальные крылья, которыми никогда не пользуются.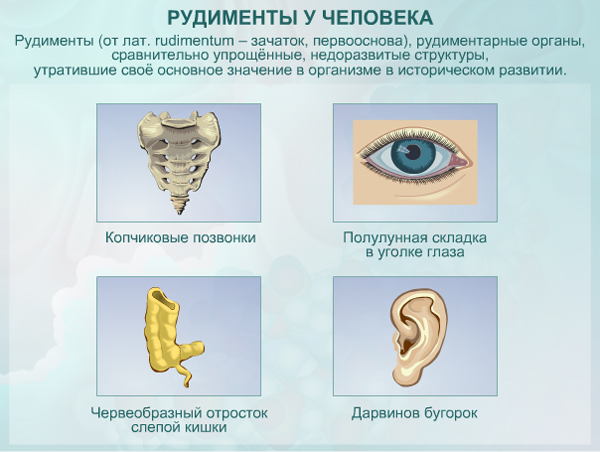 «
«
Доказательства эволюции 11 класс онлайн-подготовка на Ростелеком Лицей
Доказательства эволюции
1. Переходные формы
2. Палеонтологические ряды
3. Морфологическое доказательство
4. Эмбриологическое доказательство
5. Биогеологическое доказательство
6. Генетическое доказательство
Переходная форма
Переходная форма – организм, обладающий признаками обоих таксонов, отражающий постепенный переход от одного биологического вида к другому.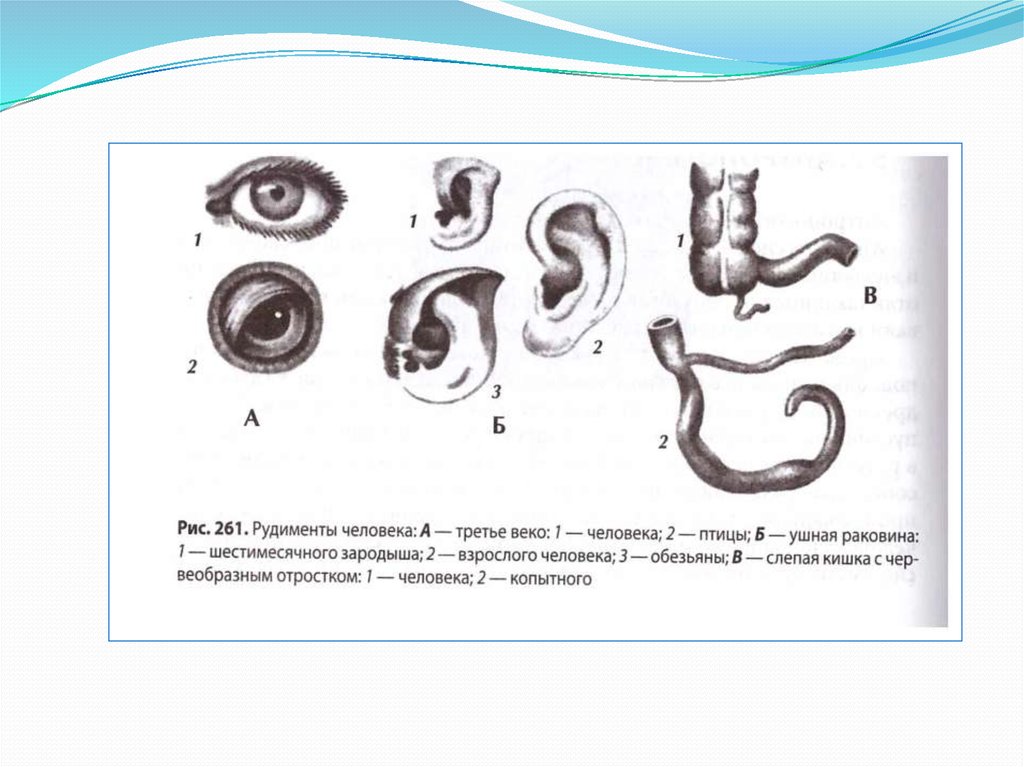
Рис. 1. Переходные формы между таксонами живых существ: археоптерикс (рептилии – птицы), семенной папоротник (папоротники – голосеменные), двоякодышащая рыба (костные рыбы – амфибии).
Существуют виды или палеонтологические останки видов, которые обьединяют черты древних и более современных организмов. Например, археоптерикс имел признаки рептилий и птиц. Семенные папоротники имели признаки папоротников и голосеменных растений. Двоякодышащие рыбы – признаки рыб и амфибий (Рис. 1).
Переходные формы считаются доказательством того, что ароморфозы происходили не внезапно, а постепенно.
Палеонтологические ряды
Палеонтологические ряды – ряды ископаемых родственных форм, которые отражают ход эволюции.
Например, палеонтологический ряд лошади показывает, как развивалась нога лошади от однопалой к пятипалой (см. видео). Копыта лошади – это роговые наросты средних пальцев конечностей.
Морфологическое доказательство
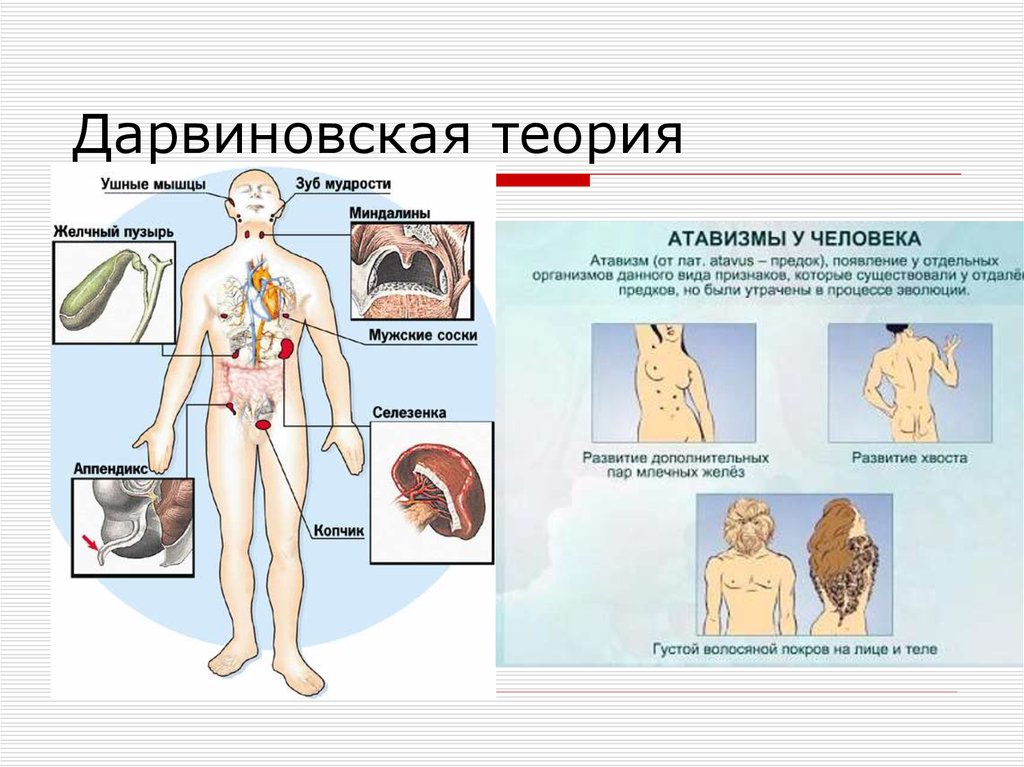
Морфологическое доказательство заключается в наличии аналогичных и гомологичных органов, рудиментов и атавизмов (рис.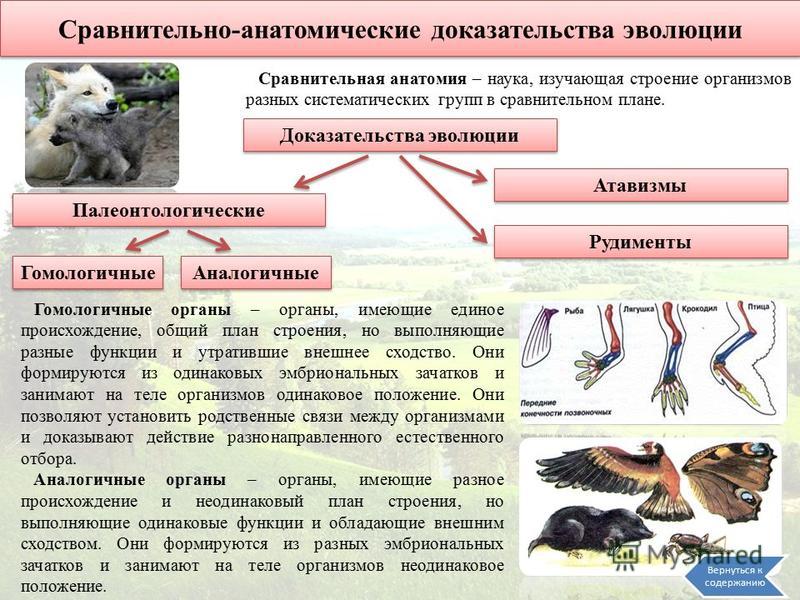 2).
2).
Гомологичные и аналогичные органы появились в процессе важнейших эволюционных процессов – конвергенции и дивергенции признаков.
Рис. 2. Морфологическое доказательство эволюции
гомологичные органы (развиваются из одних зачатков, выполняют разные функции) и аналогичные органы (развиваются из разных зачатков, выполняют оду и ту же функцию)
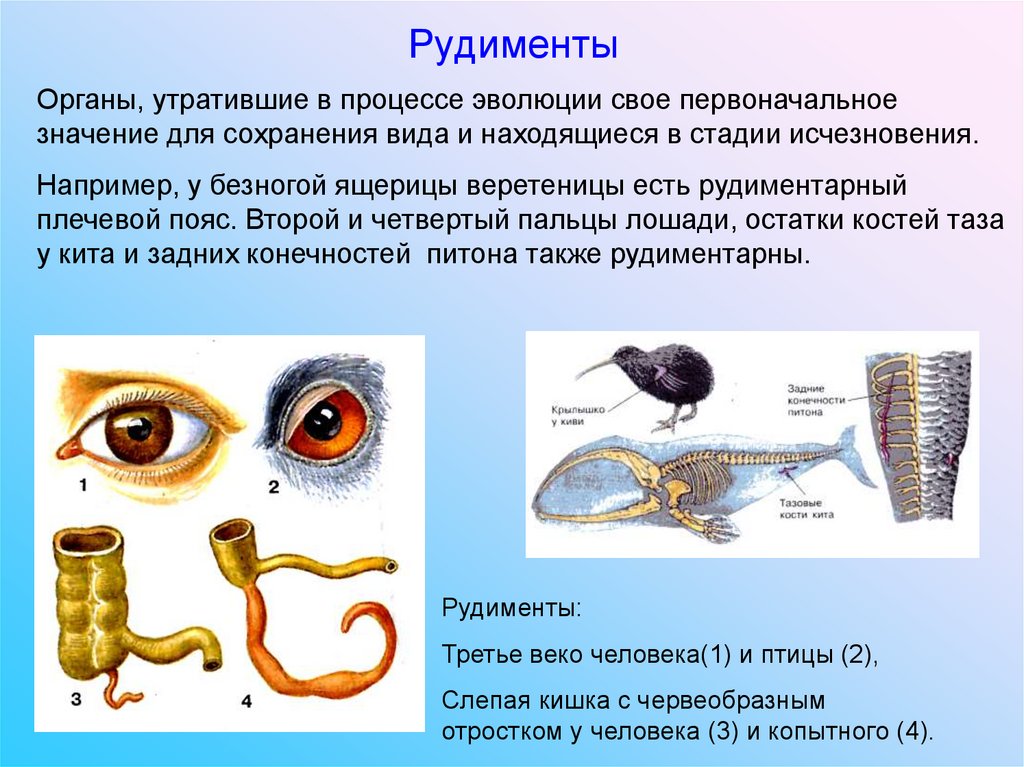
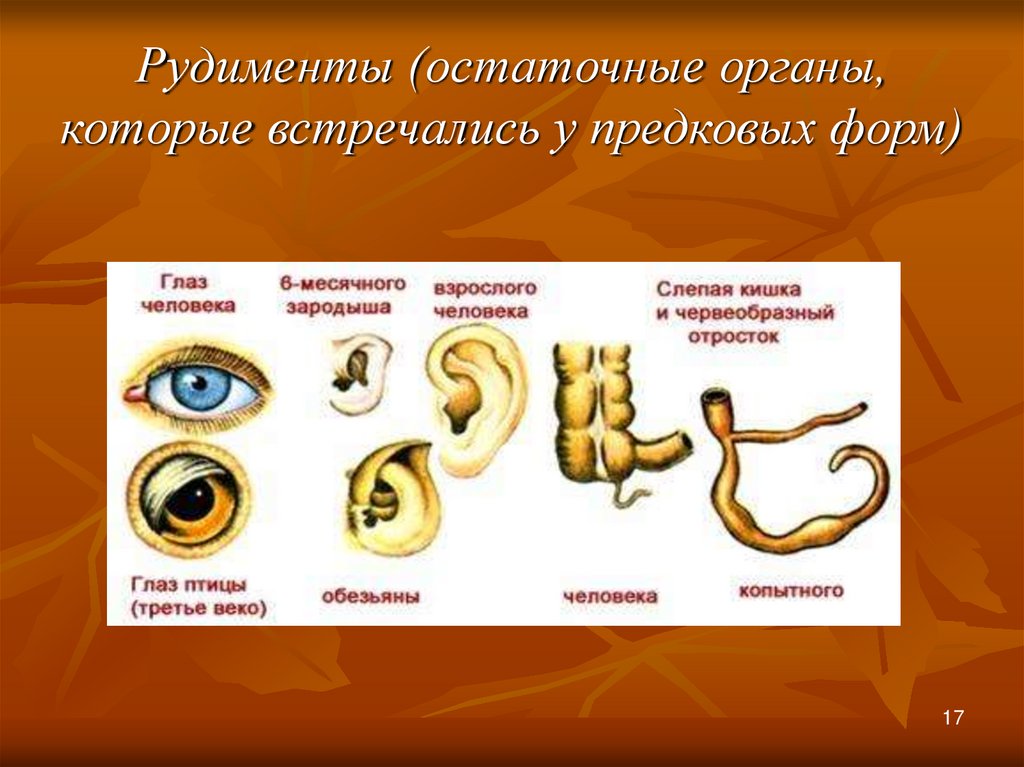
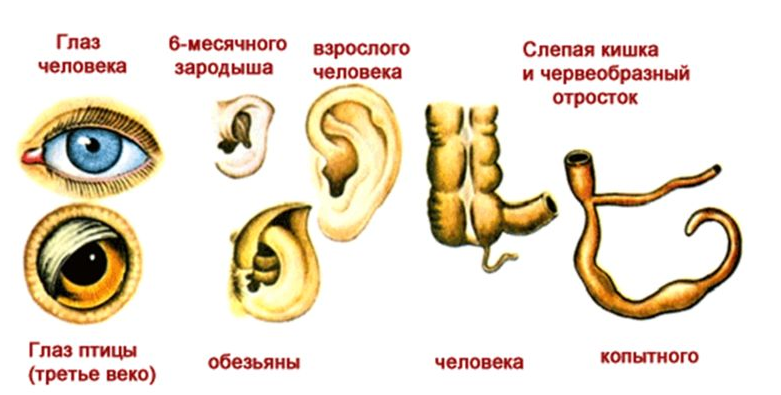
Рудименты
Рудименты – это нефункциональные (или уменьшившиеся в размерах) остатки органов живых существ, которые активно использовались и были более развиты у их предков.
Например, у китов сохранился небольшой рудимент задних ног. Это означает, что предки китов когда-то ходили на ногах, но в ходе эволюции стали водоплавающими животными (см. видео).
Атавизмы
Атавизмы – это появление у особи какого-то признака ее предка. Причем в общем случае этот признак никогда не проявляется, но у некоторых особей он может появиться. То есть, атавизм – это всегда уродство, патология.
Например, у некоторых людей развивается густой волосяной покров, как у древнего предка-примата, или многососковость, как у еще более древних млекопитающих (см.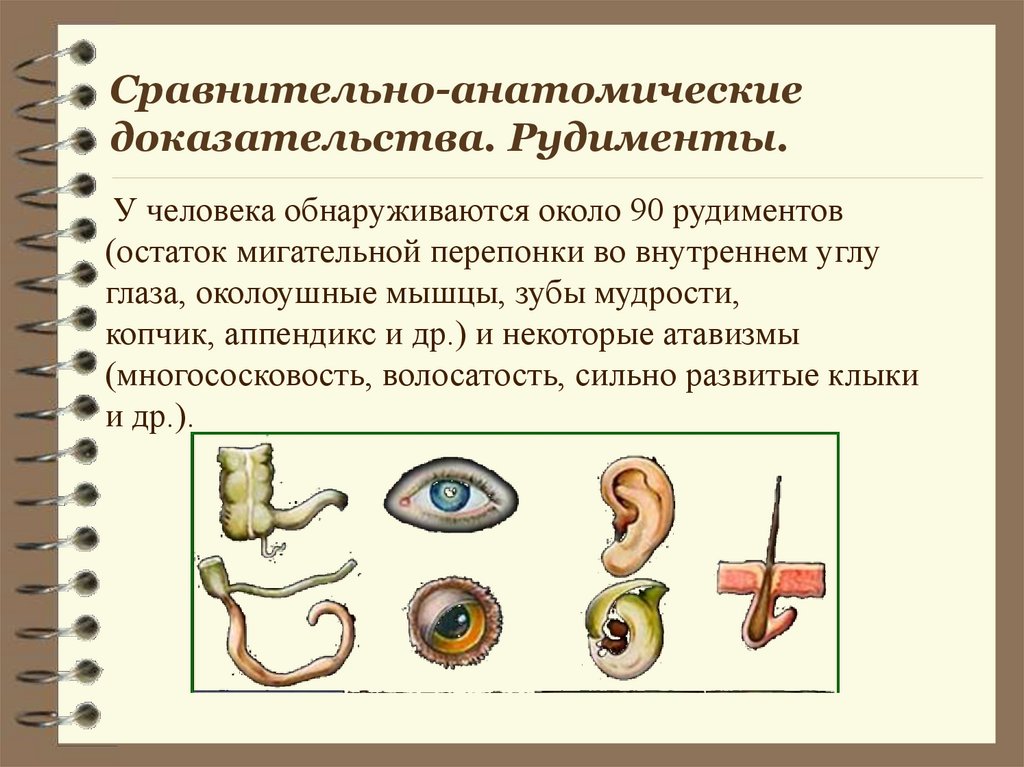 видео).
видео).
Рудименты и атавизмы доказывают, что признаки предков, так или иначе, сохраняются у потомков. Почему же они так проявляются? Ответ на это вопрос не так прост – читайте об этом в разделе Генетические доказательства.
Эмбриологическое доказательство
Эмбриологическое доказательство. Русский ученый Карл Бэр показал, что на ранних этапах онтогенеза зародыши разных видов похожи, а различия накапливаются на более поздних этапах.
О подобных вещах говорил Эрнест Геккель, формулируя биологический закон: «онтогенез есть ускоренное повторение филогенеза».
Закон Бэра
Закон Бэра: чем более ранние стадии индивидуального развитии исследуется, тем больше сходства обнаруживается между различными организмами.
Это правило можно трактовать так, что все живое развивалось из общего предка в процессе дивергенции видов.
Биогеографическое доказательство
Биогеографическое доказательство. Изучая морфологию и генетику близких видов, населяющие разделенные ареалы, можно доказать важность изоляции видообразования.
Например, своеобразие флоры и фауны Австралии обьясняется длительной изоляцией и отсутствием потока генов.
Генетическое доказательство
Генетическое.доказательство. Изучение нуклеотидных последовательностей ДНК позволило доказать наличие эволюции на молекулярном уровне.
Все эти доказательства подтверждают правоту эволюционной теории.
Список литературы
- А.А. Каменский, Е.А. Криксунов, В.В. Пасечник. Общая биология, 10–11 класс. – М.: Дрофа, 2005. По ссылке скачать учебник (Источник)
- Д.К. Беляев. Биология 10–11 класс. Общая биология. Базовый уровень. – 11-е издание, стереотипное. – М.: Просвещение, 2012. – 304 с. (Источник).
- В.Б. Захаров, С.Г. Мамонтов, Н.И. Сонин, Е.Т. Захарова. Биология 11 класс. Общая биология. Профильный уровень. – 5-е издание, стереотипное. – М.: Дрофа, 2010. – 388 с. (Источник)
- В.И. Сивоглазов, И.Б. Агафонова, Е.Т. Захарова. Биология 10–11 класс.
 Общая биология. Базовый уровень. – 6-е издание, дополненное. – М.: Дрофа, 2010. – 384 с. (Источник)
Общая биология. Базовый уровень. – 6-е издание, дополненное. – М.: Дрофа, 2010. – 384 с. (Источник)
 Общая биология. Базовый уровень. – 6-е издание, дополненное. – М.: Дрофа, 2010. – 384 с. (Источник)
Общая биология. Базовый уровень. – 6-е издание, дополненное. – М.: Дрофа, 2010. – 384 с. (Источник)
Дополнительные рекомендованные ссылки на ресурсы сети Интернет
- Ru.wikipedia.org (Источник).
- Bioword.narod.ru (Источник).
- Sbio.info (Источник).
- Bio.fizteh.ru (Источник).
- Bioenc.ru (Источник).
- Biologiya-uroki.ru (Источник).
Домашнее задание
- Почему эволюция нуждается в доказательстве? Нужны ли доказательства креационизму?
- Что вы понимаете под термином «эволюция»?
- Приведите примеры доказательства эволюционных изменений живых организмов.
- Что понимают в биологии под переходными формами?
- В чем разница между аналогичными и гомологичными органами?
- Приведите примеры рудиментов и атавизмов, которые встречаются у человека.
- Почему перечисленные на уроке факты доказывают существование эволюционного процесса?
- Обсудите с друзьями и родственниками достоверность и достаточность доказательств эволюции.

Определение сходства: историческая, биологическая и генеративная гомология.
- DOI:10.1002/1521-1878(200009)22:9<846::AID-BIES10>3.0.CO;2-R
title={Определение одинаковости: историческая, биологическая и генеративная гомология.},
автор = {Энн Б. Батлер и Уильям М. Сайдел},
journal={BioEssays: новости и обзоры молекулярной, клеточной биологии и биологии развития},
год = {2000},
объем={22 9},
страницы={
846-53
}
}- A. Butler, W. Saidel
- Опубликовано 1 сентября 2000 г.
- Biology
- BioEssays: новости и обзоры молекулярной, клеточной биологии и биологии развития эволюции и генеративных путей. Филогенетическая гомология фокусируется на происхождении признака от общего предка. Биологическая гомология касается эволюции и разнообразия признаков. Исключения из общего случая усложняют эти два подхода: исторически и биологически гомологичные признаки могут быть произведены разными генеративными путями, а признаки очень похожи. ..
Посмотреть в PubMed
doi.org
Гомоплазия и гомология: дихотомия или континуум?
Филогенез и эво-дево: признаки, гомология и исторический анализ эволюции развития.
- Дж. Кракрафт
Биология
Зоология
- 2005
Происхождение с модификацией: единство, лежащее в основе гомологии и гомоплазии, как видно из анализа развития и эволюции
7 признаков, обычно сгруппированных как гомопластические, причем последними являются конвергенция, параллелизм, инверсия, рудименты, рудименты, атавизмы и атавизмы, при этом конвергенция является единственным классом гомоплазии.
HOMOLOGY AND HOMOPLASY
- B. Hall
Biology
- 2007
How to Compare Homology Concepts: Class Reasoning About Evolution and Morphology in Phylogenetics and Developmental Biology
- M. MacLeod
Biology
- 2011
В этой статье утверждается, что движения в этом направлении упускают существенное несоответствие между этими концепциями, поскольку они были разработаны в контексте дебатов о наилучшей концепции для эволюционного исследования, и должны видеть, что эти концепции используют общую фундаментальную методологическую основу.
подход к гомологии, но не согласны с тем, как эффективно применять методологию.Полевая гомология как способ согласования генетической изменчивости и изменчивости развития с гомологией взрослых
Эво-дево и поиск гомологии («сходства») в биологических системах
В следующем эссе делается акцент на эвристических преимуществах, казалось бы, противоположных точек зрения на структурная биология, с примерами в основном из сравнительной морфологии растений.
Глубокая гомология в эпоху секвенирования нового поколения высокопроизводительное секвенирование следующего поколения.
Гомология процесса: динамика развития в сравнительной биологии
- J. Difrisco, J. Jaeger
Биология
Focus Focus
- 2021
. в частности, на примерах сегментации насекомых и сомитогенеза позвоночных, и предлагает шесть критериев гомологии процессов, сочетая признанные индикаторы с новыми, полученными в результате моделирования динамических систем.
1,05-Полевые гомологии
- L. Medina
Биология
- 2007
Показ 1-10 из 43 ссылок
Сорт-бирелевой экипаж и концепция.
- G. Striedter, R. Northcutt
Биология
Мозг, поведение и эволюция
- 1991
и на неспособности признать иерархическую природу биологической организации.
ПРОИСХОЖДЕНИЕ МОРФОЛОГИЧЕСКИХ ПРИЗНАКОВ И БИОЛОГИЧЕСКИЕ ОСНОВЫ ГОМОЛОГИИ
Развитие и изменчивость крючков грудных плавников у собачковых рыб представлены как модель для изучения простой онтогенетической сети.
Гомоплазия: результат естественного отбора или свидетельство ограничений дизайна?
Чтобы объяснить, как морфология эволюционирует в линиях, необходимы как функционалистский, так и структуралистский подходы, объединенные в контексте, в котором постоянно проводятся филогенетические гипотезы и их проверки.
Гомология, Hox-гены и интеграция развития
- Г. Мюллер, Г. Вагнер
Биология
- 1996
предполагает, что новые гомологи могут возникать в эволюции как следствие изменения эпигенетического контекста законсервированной функции генов.
Концепция биологической гомологии
- Г. Вагнер
Биология
- 1989
В этой главе рассматриваются открытые биологические вопросы, связанные с концепцией гомологии, и различные попытки установить биологическую основу гомологии в сравнительной анатомии.
Генетика развития и традиционная гомология.
- Дж. Болкер, Р. Рафф
Биология
BioEssays: новости и обзоры молекулярной, клеточной биологии и биологии развития
- 1996
Утверждается, что, хотя генетические данные о развитии могут помочь идентифицировать гомологичные структуры, они не являются ни необходимыми, ни достаточными, и ни в коем случае не оправдывают новое определение гомологии.
Молекулы и морфология: где гомология?
Гомология: иерархическая основа сравнительной биологии
- Б. Холл
Биология
- 1994
Вагнер и механизмы развития, Г.П. Шубин и М. Дж. Сандерсон одними из первых предложили концепцию «гомологии как клеточного автомата».
Гомология — история понятия.
Концепция последовательной гомологии возникла в конце 18-го века, хотя сравнение между организмами стало популярным вскоре после этого и было поддержано знаменитыми дебатами Кювье и Жоффруа Сент-Илера в 1830-х годах.
Гомология и причины.
- Л. В. Вален
Биология
- 1982
Все другие гипотезы исторической гомологии, основанные на повторяющейся гомологии, такие как лиственная теория цветка, рассматриваемого филетически, вызывают подозрение.
Истинные рудиментарные структуры у китов и дельфинов
Что такое рудиментарные структуры?
Третий новый международный словарь Вебстера определяет рудимент как «небольшой и
дегенеративная или несовершенно развитая часть тела или орган, который остался от одного
более полно развиты на более ранней стадии личности, в прошлом
поколения или в близкородственных формах». Это те рудиментарные органы, которые показывают
признаки прихода от прошлых поколений, которые поддерживают теорию эволюции.С самого начала креационисты оспаривали либо существование, либо
значение рудиментарных органов. Роберт Кофаль заявляет в своем Handy Dandy
Эволюция Опровержение :Развитие знаний и физиологии показало, что большинство предполагаемых рудиментарных
органы полезны и даже необходимы. Если есть настоящие рудиментарные
органов, они показывают потерю структуры и дизайна, а не производство
что-то новое. Но чтобы поддержать теорию эволюции, доказательства производства
требуются новые органы.С другой стороны, ведущие ученые, такие как покойный зоолог и
генетик Феодосий Добжанский, продолжали поддерживать рудиментарные органы как
доказательства эволюции. В своем тексте « Эволюция, генетика и человек » Добржанский
писал: «Действительно, нет сомнения, что рудиментарные рудиментарные органы безмолвно
провозгласить факт эволюции».Тогда, кажется, уместно говорить о заявлении Кофаля. Возьмем его
высказывание по одному предложению за раз.Когда Кофаль говорит, что развитие знаний открыло применение большинству того, что
считались рудиментарными органами, он, по крайней мере, частично прав. По сравнению с
то, что думали ученые в прошлом веке, ученые сегодня меньше обращают
органы как действительно рудиментарные. Среди недавних ученых, отметивших использование
за ранее проведенными остатками является русский зоолог Алексей Яблоков. Его книга,
Изменчивость млекопитающих , обсуждает важные функции, выполняемые тазовым
кости и усы китов — две особенности, которые раньше считались
пережитки.— стр. 9 —
Означает ли это, что по прошествии достаточного времени ученые скоро найдут применение
все так называемые рудиментарные органы, и что сегодня нам было бы благоразумно дать
выдвигать аргумент об рудиментарных органах как обреченный случай? Это то, что
креационисты хотели бы, чтобы мы предположили. Но такое отношение основано на
непонимание того, почему многие структуры, считавшиеся в прошлом пережитками,
сегодня считается чем-то другим.В главе, озаглавленной «Изменчивость и проблема рудиментарных органов»,
Яблоков помогает нам разобраться в недоразумении. Он видит большую часть проблемы как
были вызваны «расплывчатыми определениями концепции рудиментарных органов»,
особенно «расплывчатое или неточное понимание концепции рудиментарного
орган, присутствующий в трудах Дарвина…» (стр. 232-233). Его цель состоит в том, чтобы
уточнить определения, чтобы дать ученым более четкое представление о
какие критерии должны быть соблюдены, прежде чем структура может быть названа рудиментарной.Но даже при более строгих определениях органы, которые больше не помечаются как рудиментарные,
не перестают иметь значение для демонстрации эволюции, как могли бы подумать креационисты.
Например, когда Яблоков отрицает, что таз и усы у китов действительно
рудиментарными, он продолжает утверждать, что они являются явным возвратом к более раннему
эволюционный этап. Как он утверждает на странице 240, «структура этих органов была
модифицированы значительным изменением функции в какой-то момент их эволюции».
Из-за своих более жестких определений он предпочитает говорить в таких случаях «о
рудиментарность функций, а не рудиментарность органов» (стр. 246).
означает, что креационисты очень мало выигрывают от «продвинутого знания», которое поддерживает
обнаружение полезных, но измененных функций органов, ранее определявшихся как
«рудиментарный». Эти органы все еще демонстрируют происхождение с модификацией.По мнению Яблокова, проблема с большинством ранее существовавших примеров
рудиментарными органами состоит в том, что они были органами, «присутствующими у всех особей
данного вида». Но «во всех случаях наблюдается, что такие органы или
структуры, унаследованные всем населением, имеют функциональное значение
и логически не может быть назван рудиментарным». Для Яблокова орган должен быть
считается рудиментарным только в том случае, если оно развивается у некоторых индивидуумов, но
не свойственна всему населению (с. 241). И добавляет: «Было
давно известно, что такие органы существуют у животных».0021Вооружившись этим более четким представлением о рудиментарных органах, мы можем теперь взглянуть на
второе предложение в заявлении Кофала. Здесь он говорит, что всякое истинное рудиментарное
орган показал бы потерю структуры и дизайна вместо развития
что-то новое. Если мы отбросим слово «дизайн», Кофаль совершенно прав. Правда
Рудиментарный орган — это действительно орган, утративший свою первоначальную структуру. Оно имеет
также потерял свою функцию. Лица, обладающие рудиментарным органом, не отличаются
фитнес от тех, у кого его нет. Орган просто остаток. Следовательно, Кофаль
кажется, принимает концепцию.Но когда он говорит о «дизайне», он подразумевает дизайнера, мастера-архитектора, который
ставит органы на место; животные были идеально созданы совершенным мастером, который
точно знал, что нужно делать.— стр. 10 —
Учитывая такое положение, казалось бы
легкомысленно, если этот производитель поместил некоторые части, которые были бесполезны. Но «дизайн» — это
концепция, которая не имеет никакой полезности в науке. Как указывает Ричард Оли:Мы не наблюдаем дизайна в природе. Скорее наши умы, кажется, устроены так
что мы можем воспринимать закономерности, к которым, если у нас есть религиозные
предпосылки, мы применяем концепцию дизайна. Кроме того, чтобы сделать дизайн
а
биологический принцип. . . заключается в уменьшении необходимости интерпретировать биологические процессы
как предшественники адаптации, вызывающей удивление.Это подводит нас к последнему предложению Кофаля. Там он утверждает, что, поскольку рудиментарное
органы демонстрируют потерю структуры, они не помогают эволюции, поскольку эволюция
требует «доказательств производства новых органов…». Это предложение показывает
Непонимание Кофалом того, почему рудиментарные органы используются в качестве доказательства в
поддержка эволюции.Рудиментарные остатки восходят к более ранней эволюционной стадии, потому что они
рудименты – это органы или части, которые перестали выполнять свои первоначальные функции.
функционировать и стали ненужными и атрофировались. Яблоков считает их
примеры атавизма, «органов, появляющихся в развитии настоящих форм и
указывающее на состояние их предков (atavus = предок на латыни)» (стр.
244). Производство новых органов не имеет к этому никакого отношения. Это
совершенно отдельная эволюционная проблема.Появление конечностей у китообразных
Теперь, когда у нас есть более полное представление о том, что представляет собой настоящий рудиментарный
структуру, мы можем перейти к поиску конкретного примера. Китообразные (киты
и дельфины) обычно считаются обладающими многими пережитками. Конечно, это
именно эти млекопитающие использовались Яблоковым для демонстрации
ошибки, допущенные учеными в отношении рудиментарных структур. Поэтому мы должны спросить,
Находит ли Яблоков какие-либо доказательства существования истинных рудиментарных структур у китообразных?
Да, знает, и большая часть его данных основана на личном расследовании.Среди рудиментарных структур китообразных, которые он принимает, есть рудиментарные задние.
конечности. Ему известно о шести случаях только у кашалотов. Особенно один, который
позже был передан ему для личного изучения, он обсуждает на странице 242.В июне 1962 г. В. 1. Борисов наблюдал кашалота с хорошо развитыми
выпуклости на брюшной части тела при работе в ките
завод в Скалистии (Средние Курилы). Один из этих выступов может
даже сделать рентген.Сам Яблоков наблюдал остаток бедренной кости у самца кашалота в
завод в Подгорном (Северные Курилы).— стр. 11 —
Другим следователям также известны доказательства этого типа. Уильям Кинг
Грегори, написав в 1962 году в Британской энциклопедии , предоставил следующее:
учетная запись.В июле 1919 года самка горбатого кита (Megaptera nodosa) с двумя замечательными
выступы на вентральной стороне тела, кзади, были захвачены кораблем
работает с китобойной станции в Кюквоте, на западном побережье Ванкувера.
Остров, Британская Колумбия. Один из выступов был срезан экипажем.
судно, а другое было сфотографировано на месте суперинтендантом
станция.По просьбе Роя Чепмена Эндрюса скелетные останки, которые состояли
из двух костей и двух тяжелых хрящей, были отправлены из Канады в американскую
Музей естественной истории в Нью-Йорке. Найденный образец имел элементарные
ноги торчат из тела примерно на четыре фута два дюйма, покрытые
с ворванью толщиной примерно в полдюйма. Эндрюс идентифицировал кости как большеберцовую кость.
и плюсневых, хрящей, таких как бедренная кость и предплюсна, и опубликовал свои выводы:Изучив материал и обсудив его с разными учеными, я
пришли к выводу, что выступы на самом деле представляют собой рудиментарные задние
конечностей и демонстрируют замечательную реверсию к примитивному четвероногому состоянию.У профессора Эндрюса было достаточно анатомических причин, чтобы отвергнуть идею о том, что
конечности были просто аномальными пороками развития без реверсионного значения. Он
завершил свое исследование этого замечательного экземпляра следующим
наблюдения:Поскольку исследования Кукенталя и Гульдберга показали, что наружная задняя конечность
зачатки еще присутствуют в некоторых случаях в зародышевой жизни, это отнюдь не
невозможно, чтобы эти рудиментарные органы продолжали свой рост и сохранялись
до взрослой стадии. Я считаю, что именно это и произошло в
образец, который я описал выше, и что мы сталкиваемся с явным
случай частичного возврата к примитивному четвероногому состоянию.Конечности, по утверждениям китобоев, были симметричны; Oни
находятся в точном положении, в котором были обнаружены зачатки задних конечностей у
эмбриональные Megaptera; есть явные признаки того, что хрящевая ткань бедренной кости
прикреплялась к тазовым элементам.В отчете, озаглавленном «Замечательный случай наружных задних конечностей у горбатого
Кит», был опубликован в июне 1921 года.В 1953 году Тейдзо Огава написал «О присутствии и исчезновении лань».
Конечности в эмбрионах китообразных» в Научный отчет об исследованиях китов ,
пришел к выводу, что:— стр. 12 —
… на мой взгляд, исчезновение парных задних конечностей у китообразных
кажется, имеет тесную причинно-следственную связь с появлением парного каудального
флюсы из них. . . . У эмбриона дельфина Prodelphinus длиной 14 мм.
caeruleoalbus, а у зародыша горбача Megaptera nodosa длиной 20 мм
парные возвышения задних конечностей развиты довольно хорошо. Фотографии
они показаны. Дополнительное внимание было уделено одновременности
исчезновение возвышения задних конечностей с первым появлением хвостового
сосальщиков у эмбрионов китообразных. (стр. 131)В 1957 году Тецио Огава и Тоширо Камия в том же журнале сообщили
на тему «Случай кашалота с оттопыренной рудиментарной задницей».
Конечности.»Излишне говорить, что у всех китообразных в
их послеродовая жизнь. Только на ранней эмбриональной стадии они показывают пару
оттопыренные задние конечности, которые вскоре исчезают. С другой стороны, существование
пары мелких тазовых костей известны почти всем китообразным, лежащим
далеко от позвоночного столба по обе стороны от полового отверстия. в
у финных и синих китов, а у горбатых бедренная кость тоже имеется у
таз, а у кита есть даже большеберцовая кость. Конечно, эти кости
глубоко погружается под кожу, не образуя выпуклостей на поверхности тела. (п.
197)После некоторого обсуждения отчета Эндрюса 1921 года авторы продолжили:
Однако недавно другая особь, принадлежащая Odontoceti [кашалоту],
и обладающий также парой оттопыренных задних конечностей, встречался у
Япония… Согласно отчету, представленному с китобойной станции, это был
самка длиной 10,6 м. Выпуклости были с обеих сторон
полового отверстия…. (стр. 198)Различие между двумя случаями никогда не является существенным, а скорее является проблемой
количество материалов для изучения.В своем резюме Огава и Камия заявили:
У почти взрослой самки кашалота, пойманной в ноябре 1956 года у Кинквазана в
Япония, присутствовала пара рудиментарных задних конечностей, похожих на почки. Высота
выпуклость справа 5,35 см, слева 6,56 см. На
осмотр внутренней части левой конечности, трех частично хрящевых костей
были найдены. Они соответствовали тазу, бедру и, возможно, большеберцовой кости, но не
между ними есть стыки. Довольно сильные мышцы соединяют таз и
бедро, а между бедром и большеберцовой костью простираются две слабые мышцы… .Этот случай можно понять, если предположить аномальное сохранение ранней
эмбриональном состоянии и показывает, весьма вероятно, атавизм обратно к четвероногому
состояние далеких предков китов. Это никогда не может быть пороком развития, не имеющим филогенетического значения. (стр. 207)— стр. 13 —
Эти примеры рудиментарных задних конечностей соответствуют критерию Яблокова.
нефункциональными, а также нехарактерными для всего населения. К
демонстрируют их нехарактерный характер, доклад Сэйдзи Осуми
поучительный.16 декабря 1963 года стадо из примерно 450 сине-белых дельфинов (Stenella
caeruleoalba) был пойман рыбаками на пляже Кавана на восточном побережье.
полуострова Идзу, Япония. В ходе биологического исследования стада
Я нашел особь с оттопыренными рудиментарными задними конечностями… оттопыренными
по обе стороны только от щели молочной железы. (стр. 135)Из 450 он нашел только один пример.
Можно было бы привести и другие рудиментарные сообщения о морских млекопитающих, но уже достаточно
представлены, чтобы показать, что эти рудиментарные задние конечности существуют у китообразных и
действительно рудиментарны. Будучи рудиментарными, они указывают на более раннюю стадию
эволюционное развитие. Из-за этого и других доказательств Уильям Кинг
Григорий заключил в своем Британская энциклопедия статья, «что у китообразных есть
был получен из наземных четвероногих плацентарных».Само слово «рудиментарный» происходит от латинского vestigium, что означает «шаг» или «шаг».
отслеживать. Рудиментарные органы — это следы ранее функционировавших органов. В некотором смысле,
рудиментарные останки подобны следам, уводящим нас в прошлое, когда
они были полностью развиты, время, когда предковое животное имело значительно
другое строение тела и совершенно другой образ жизни от примера
жив сегодня.
 ..
..