Содержание
Генетики «редактируют» родословное древо птиц
: 30 Мар 2015 , Наша Арктика , том 61,
№1
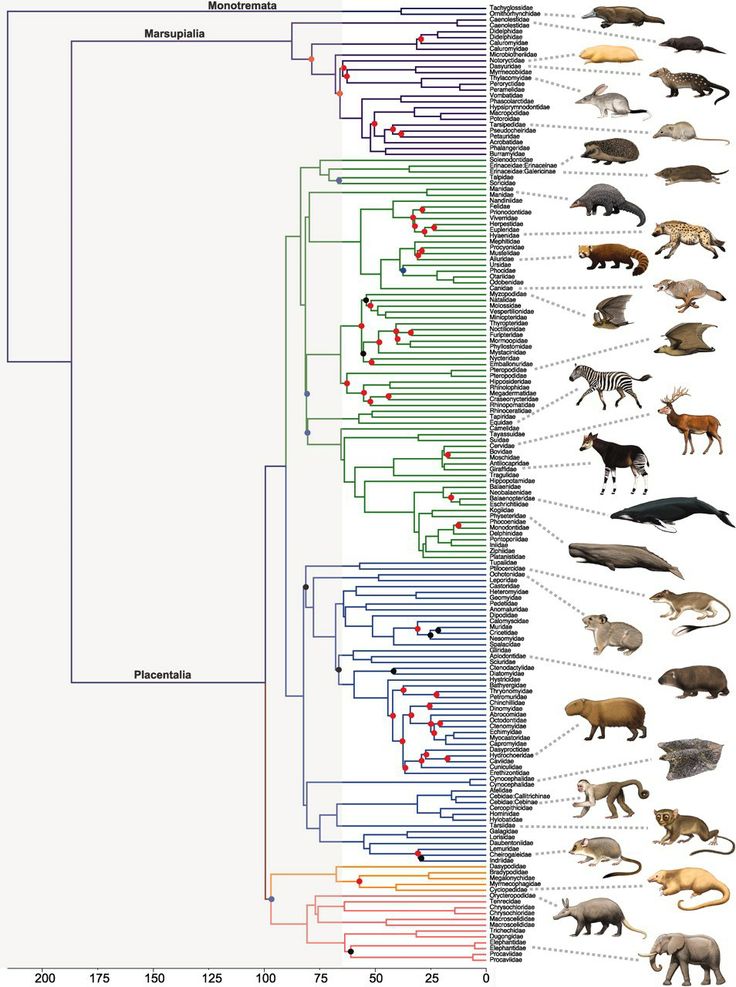
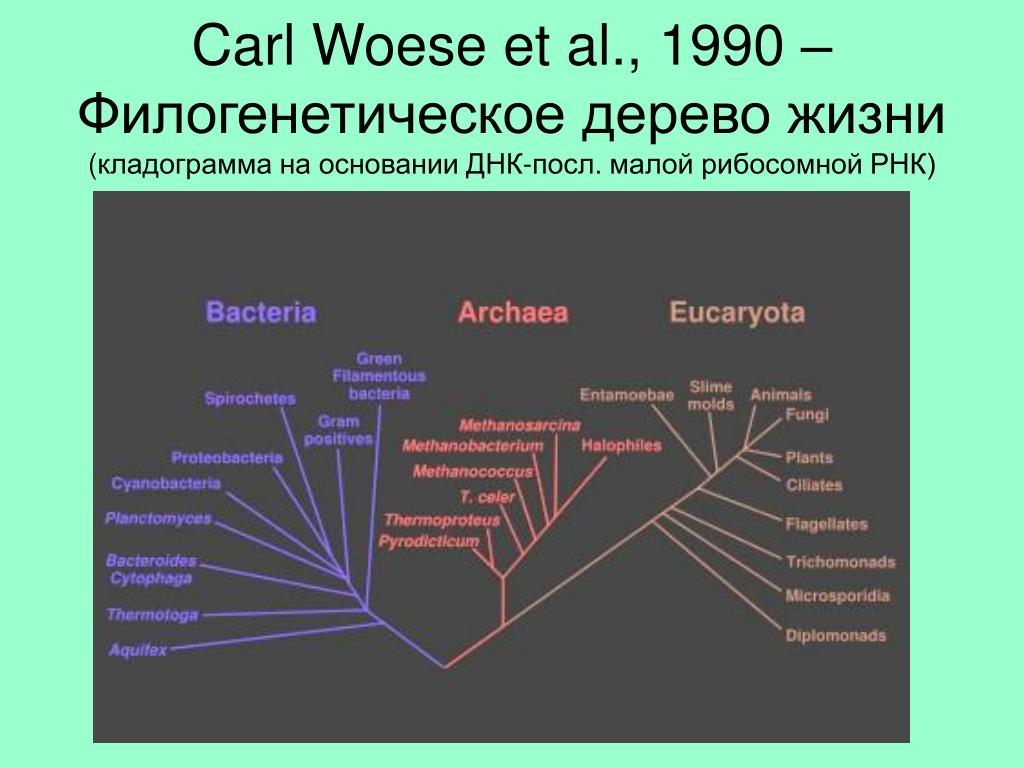
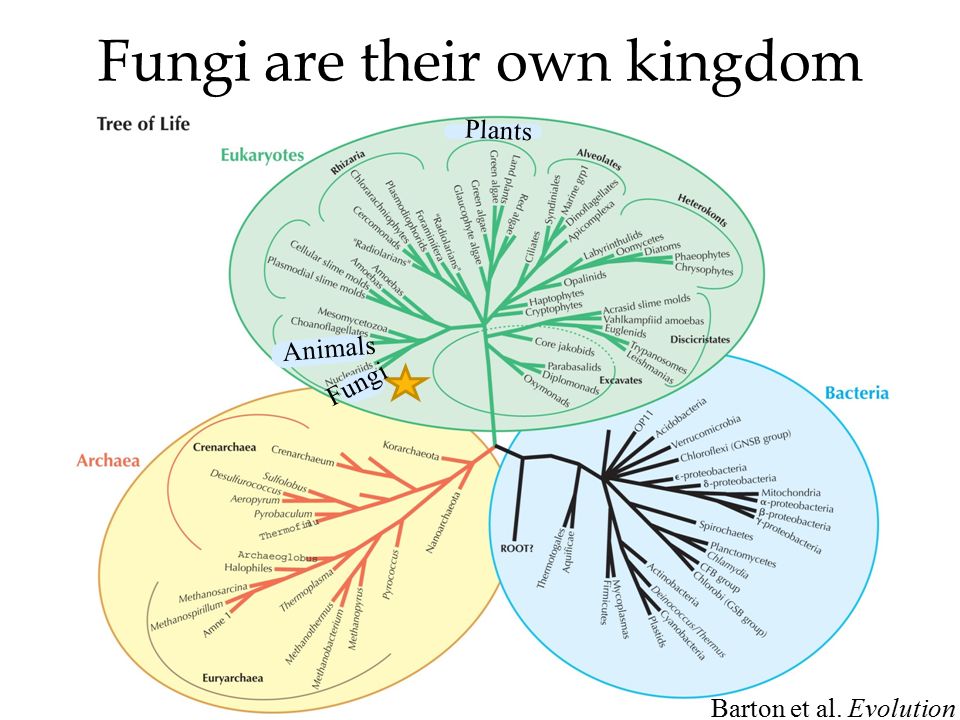
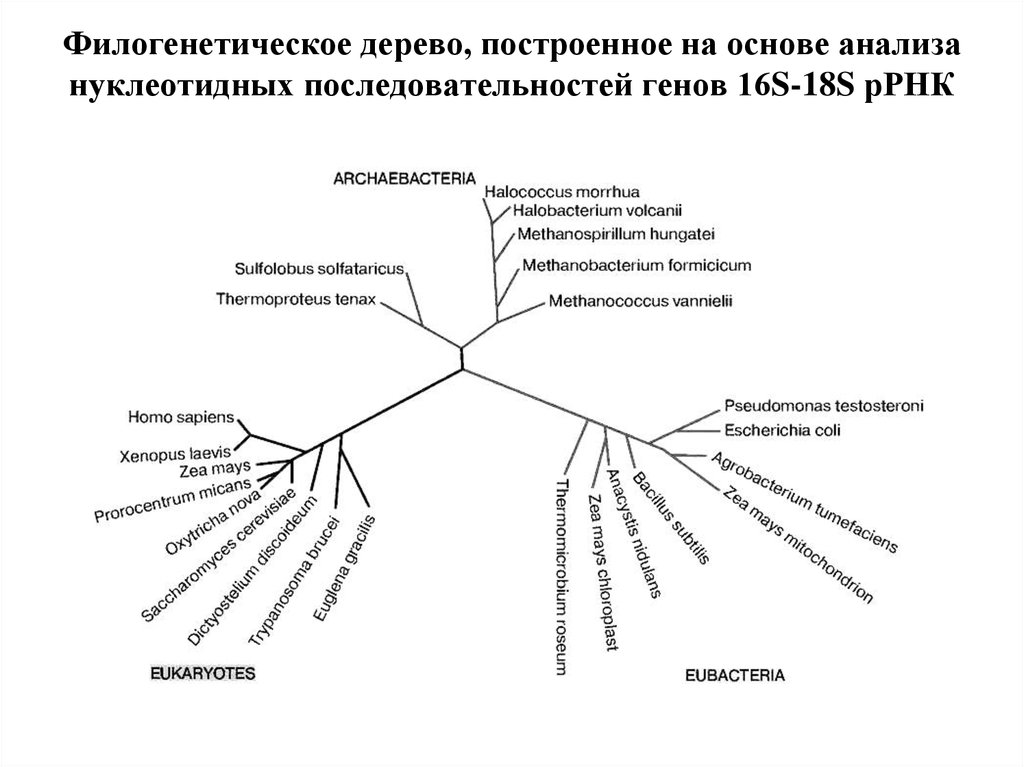
С момента зарождения теории эволюции Дарвина ученые пытаются воссоздать древо жизни для всех ныне живущих на земле организмов, и именно птицы долгое время «задавали тон» в подобных исследованиях. Однако древо, отражающее представления ученых конца XIX в. о родственных отношениях птиц, мало менялось в течение последующих десятилетий, а в некоторых странах (например, в России) орнитологи предпочитают придерживаться его и поныне. Ситуация в корне переменилась в 1990 г., когда американские орнитологи Ч. Сибли и Д. Олквист построили совершенно новое филогенетическое древо птиц исключительно на основе сходства их ДНК, не оставив и следа от традиционной классификации птиц. С этого времени отмечается всевозрастающий интерес к проблеме родственных отношений между птицами. Однако до последнего времени все работы базировались на поиске сходства последовательностей нуклеотидов в молекулах ядерной ДНК. И только в 2014 г. в Science было опубликовано новое филогенетическое древо, построенное на основе исследования полного генома у представителей всех отрядов птиц. Согласно новому древу, например, ближайшими родственниками фламинго и поганок в нашей фауне могут оказаться голуби, а ближайшими родственниками дроф – кукушки. Но особенно важно то, что теперь мы знаем, что современные представления о родстве между птицами базируются на основе сходства не только между отдельными генами и их комбинациями, но и всего генома
Однако до последнего времени все работы базировались на поиске сходства последовательностей нуклеотидов в молекулах ядерной ДНК. И только в 2014 г. в Science было опубликовано новое филогенетическое древо, построенное на основе исследования полного генома у представителей всех отрядов птиц. Согласно новому древу, например, ближайшими родственниками фламинго и поганок в нашей фауне могут оказаться голуби, а ближайшими родственниками дроф – кукушки. Но особенно важно то, что теперь мы знаем, что современные представления о родстве между птицами базируются на основе сходства не только между отдельными генами и их комбинациями, но и всего генома
Второе место среди десятки самых значительных научных достижений 2014 г. редакторы журнала Science отдали работам большого коллектива эволюционных биологов, которые с помощью современных молекулярно-генетических методов провели «ревизию» родословной птиц – наиболее многочисленного и широко распространенного класса современных позвоночных, – оценив темп и направление эволюционных переходов внутри этой группы на основе анализа геномов представителей разных таксонов пернатых
Птицы на протяжении столетий привлекали особое внимание исследователей, поэтому неудивительно, что многие открытия в различных областях биологической науки были сделаны при исследовании именно этих высших позвоночных.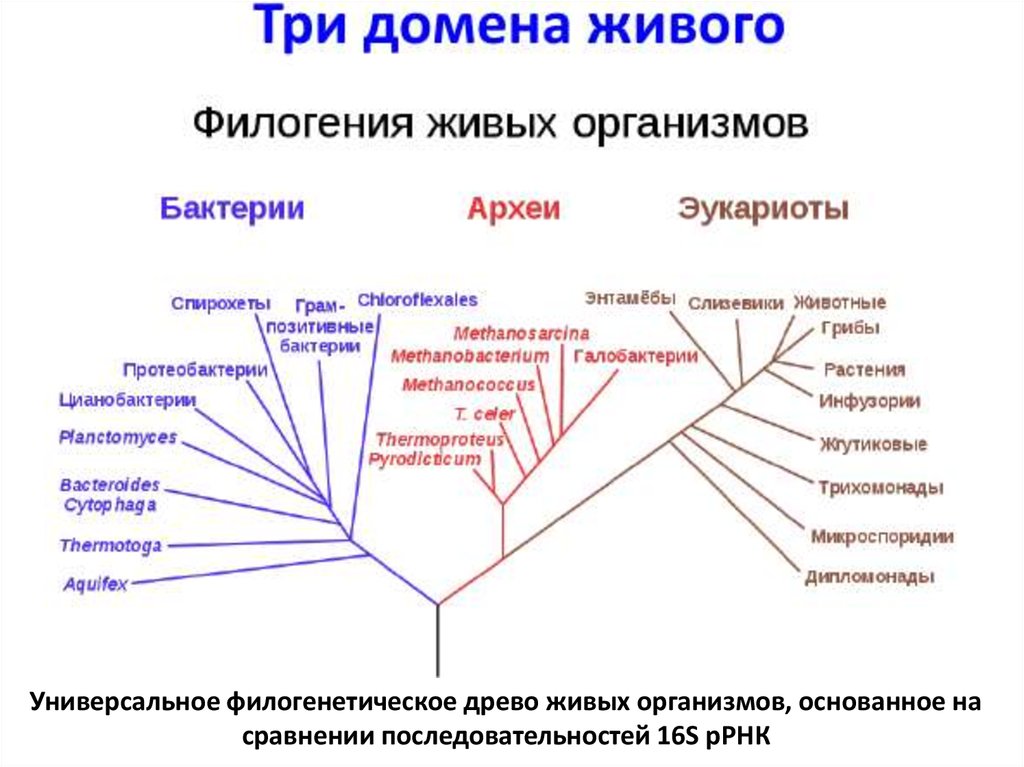 Вспомним, что даже в формировании теории происхождения видов важнейшую роль сыграли галапагосские вьюрки, так что этих птиц теперь по праву называют «дарвиновыми». Эти небольшие певчие птицы, относящиеся к танагровым – семейству всеядных птиц Нового Света, не так давно заселили Галапагосский архипелаг, где успели «произвести» на свет более десяти видов, различающихся размером, окраской оперения и строением клюва. Во время своего знаменитого путешествия на корабле «Бигль» Чарльз Дарвин посетил Галапагосы, где и наблюдал удивительное разнообразие этих птиц, являющееся более чем наглядной и убедительной иллюстрацией их недавней эволюции (по современным оценкам, предок современных галапагосских вьюрков попал на острова не ранее 2—3 млн лет назад).
Вспомним, что даже в формировании теории происхождения видов важнейшую роль сыграли галапагосские вьюрки, так что этих птиц теперь по праву называют «дарвиновыми». Эти небольшие певчие птицы, относящиеся к танагровым – семейству всеядных птиц Нового Света, не так давно заселили Галапагосский архипелаг, где успели «произвести» на свет более десяти видов, различающихся размером, окраской оперения и строением клюва. Во время своего знаменитого путешествия на корабле «Бигль» Чарльз Дарвин посетил Галапагосы, где и наблюдал удивительное разнообразие этих птиц, являющееся более чем наглядной и убедительной иллюстрацией их недавней эволюции (по современным оценкам, предок современных галапагосских вьюрков попал на острова не ранее 2—3 млн лет назад).
Теория эволюции Дарвина оказала важнейшие влияние на развитие современной науки. Одним из существенных ее следствий стало понимание, что жизнь на Земле можно представить в виде своеобразного древа – что ныне живущие виды происходят от каких-то вымерших предков, а те, в свою очередь, восходят к каким-то еще более далеким предкам. По мере удаления в прошлое «ветви», представленные современными видами, сливаются во все более толстые «сучья» и заканчиваются единым «стволом» – предполагаемым предком всего живого на Земле. Очень примечательно, что именно такое дерево жизни, которое можно уподобить генеалогическому дереву человеческого рода, было единственной иллюстрацией в первом издании знаменитой книги Ч. Дарвина «Происхождение видов…».
По мере удаления в прошлое «ветви», представленные современными видами, сливаются во все более толстые «сучья» и заканчиваются единым «стволом» – предполагаемым предком всего живого на Земле. Очень примечательно, что именно такое дерево жизни, которое можно уподобить генеалогическому дереву человеческого рода, было единственной иллюстрацией в первом издании знаменитой книги Ч. Дарвина «Происхождение видов…».
С момента зарождения теории эволюции Дарвина ученые пытаются воссоздать древо жизни для всех ныне живущих на земле организмов, и именно птицы долгое время, что называется, «задавали тон» в подобных исследованиях. Во второй половине XIX в. материал для реконструкции такого древа жизни (или, на современном языке – «филогенетического древа») ученые черпали во внутреннем строении организмов. Уже Дарвину было понятно, что внешнее строение животных может меняться в зависимости от условий среды, при этом животные, приспосабливаясь к сходным условиям, могут стать удивительно похожими друг на друга. В этом смысле хорошим примером являются такие рыбы, как акулы, и касатки – водные млекопитающие.
В этом смысле хорошим примером являются такие рыбы, как акулы, и касатки – водные млекопитающие.
В XIX в. считалось, что анатомия внутренних органов гораздо меньше или даже совсем не зависит от условий среды, и поэтому именно она годится для выявления родственных связей между организмами. С этой целью была исследована мускулатура, кости, кровеносная система и некоторые другие системы органов различных видов птиц. На основе сходства внутреннего строения и благодаря усилиям ряда ученых к концу этого столетия удалось построить вполне сносное филогенетическое древо птиц, которое более или менее исправно служило биологам и на протяжении большей части XX в. Самую заметную роль в создании базисных идей о взаимном родстве птиц сыграл немецкий анатом М. Фюрбрингер, издавший в 1888 г. поистине гигантский фолиант по сравнительной анатомии птиц, содержавшей свыше 1700 страниц современного формата A3 (Fürbringer, 1888).
Нужно сказать, что древо, отражающее представления ученых конца XIX в.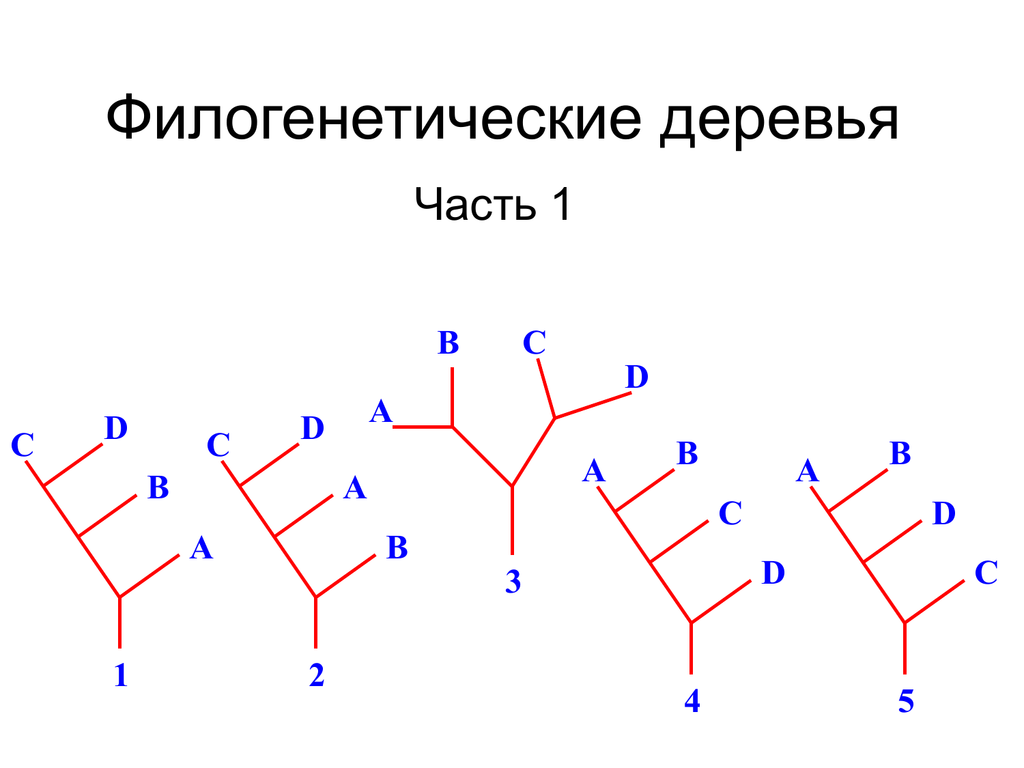 о родственных отношениях птиц, мало менялось в течение последующих десятилетий, а в некоторых странах (например, в России) орнитологи предпочитают придерживаться его и поныне. Откройте любой определитель птиц на русском языке, и вы убедитесь, что он начинается с поганок и гагар – птиц, хорошо приспособленных к водному образу жизни и поэтому похожих, но далеко не близкородственных. Хотя уже в 1970—1980-х гг. стало ясно, что в традиционных представлениях о родстве между птицами имеются многочисленные ошибки, что внутренняя анатомия также подвержена параллельной эволюции, поэтому сходство между птицами может не отражать их происхождение от недавнего общего предка.
о родственных отношениях птиц, мало менялось в течение последующих десятилетий, а в некоторых странах (например, в России) орнитологи предпочитают придерживаться его и поныне. Откройте любой определитель птиц на русском языке, и вы убедитесь, что он начинается с поганок и гагар – птиц, хорошо приспособленных к водному образу жизни и поэтому похожих, но далеко не близкородственных. Хотя уже в 1970—1980-х гг. стало ясно, что в традиционных представлениях о родстве между птицами имеются многочисленные ошибки, что внутренняя анатомия также подвержена параллельной эволюции, поэтому сходство между птицами может не отражать их происхождение от недавнего общего предка.
Одними из первых на недостатки традиционной классификации птиц обратили внимание, пожалуй, палеонтологи, изучавшие вымерших представителей. Анатомы, которые к этому времени начали исследовать внутреннее строение животных совершенно на другом уровне, также стали задаваться вопросами о родственных отношениях птиц между собой.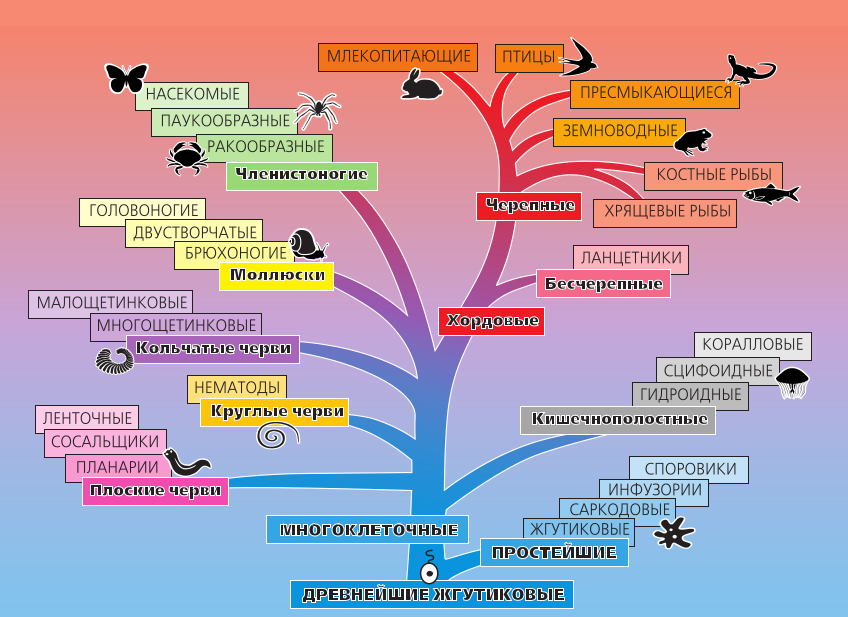 Отдельные ученые предпринимали попытки улучшить птичье филогенетическое древо, однако эти усилия не имели особого успеха – во многом потому, что орнитологическое сообщество упорно не хотело пересматривать традиционные представления о родстве пернатых между собой.
Отдельные ученые предпринимали попытки улучшить птичье филогенетическое древо, однако эти усилия не имели особого успеха – во многом потому, что орнитологическое сообщество упорно не хотело пересматривать традиционные представления о родстве пернатых между собой.
Ситуация в корне переменилась в 1990 г., когда была опубликована эпохальная книга американских орнитологов Ч. Сибли и Д. Олквиста, посвященная молекулярной эволюции птиц (Sibley, Ahlquist, 1990), – первая в истории науки работа, охватившая молекулярную эволюцию крупной группы животных. Авторы книги построили совершенно новое филогенетическое древо птиц исключительно на основе сходства их ДНК, при этом от традиционной классификации птиц не осталось и следа. Например, на удивление всех орнитологов, ближайшими родичами певчих птиц выступили голуби, журавли и аисты, но вовсе не дятлы, как это считалось ранее. Зато дятлы и туканы вдруг оказались представителями одной из самых древних независимых линий, при том что они и внешне, и по внутреннему строению очень похожи на певчих птиц.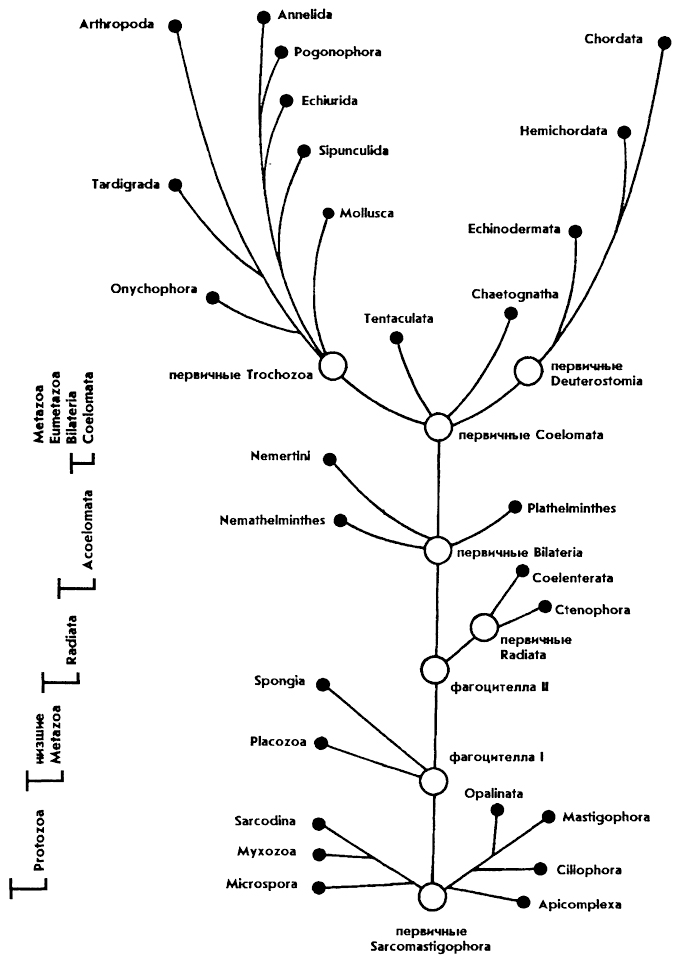
Новая классификация птиц Сибли и Олквиста была встречена очень резкой критикой – во многом из-за проблем с методологией. Время показало, что это филогенетическое древо действительно было во многом ошибочным, однако его значение для развития науки об эволюции птиц трудно переоценить. Именно эта работа заставила рядовых орнитологов допустить саму мысль о том, что традиционные представления о родственных отношениях между птицами могут быть в принципе неверными.
С середины 1990-х гг. отмечается всевозрастающий интерес к проблеме родственных отношений между птицами. По этой теме ежегодно публикуются десятки работ, благодаря чему у нас постепенно выстраивается все более ясная картина нового древа жизни птиц. Какие-то из традиционных воззрений подтверждаются, какие-то полностью отвергаются. При этом на первое место по значимости выходят данные анализа последовательности нуклеотидов в молекулах ядерной ДНК (не просто сходство, как у Сибли и Олквиста, а «продвинутое» сходство).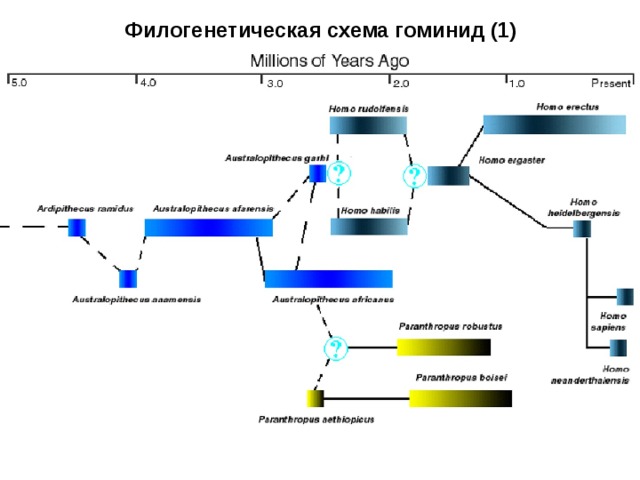
Исследования отдельных генов и редких геномных изменений позволили выявить много нового и порою неожиданного. Так, в 2001 г. было обнаружено, что ближайшими родственниками поганок являются вовсе не гагары, так на них похожие, а фламинго, с которыми у поганок, казалось бы, нет ничего общего! Поначалу сближение поганок и фламинго представлялось какой-то ошибкой молекулярных биологов, однако исследования все новых и новых генов подтверждали их близкое родство. Позже выяснилось, что у поганок и фламинго на пальцах ног имеются ногти, а у всех остальных птиц – когти, что добавило уверенности в правомерности сближения этих птиц. В конце концов удалось найти и давно вымершую группу птиц – промежуточную между фламинго и поганками. Ими оказались ископаемые фламинго палелодиды, которые в отличие от современных были очень хорошо приспособлены к плаванию и, возможно, даже к нырянию (хотя современные фламинго в принципе могут плавать, однако они мало адаптированы к такому занятию).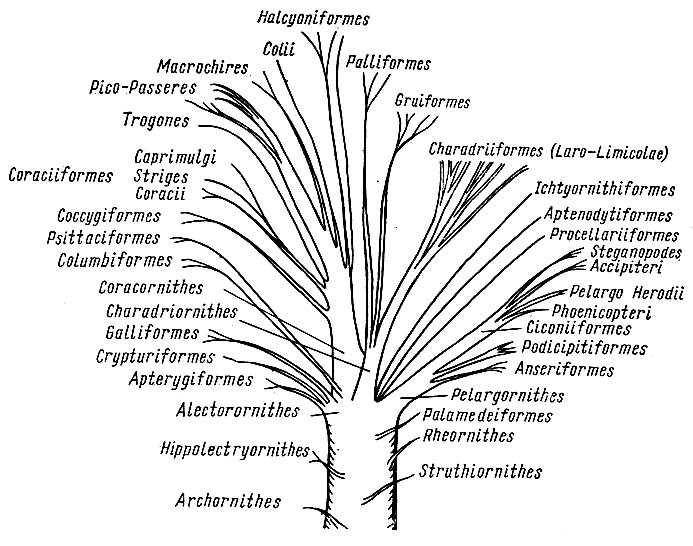
В результате тщательнейших исследований, проведенных множеством специалистов за последние 15 лет, представления о древе жизни птиц были полностью переработаны. Если бы мы сегодня создавали новый определитель птиц России, то должны были бы начинать его уже не с гагар и поганок, а с гусей, уток и фазанов, которые оказались близкими родственниками и самыми примитивными из наших птиц. Поганки и фламинго, о которых шла речь выше, могли бы идти следом. Ближайшими родственникам певчих птиц могут быть либо попугаи, либо соколы. А вот ястребы, которые раньше считались родственниками соколов, теперь помещаются совсем в другую эволюционную ветвь. Уже нет сомнений, что стрижи и колибри родственны козодоям, а цапли – не аистам, а веслоногим (например, пеликанам). Что дрофы – это отдельная линия птиц, не родственная журавлеобразным, как считалось ранее. Такую новую классификацию птиц, учитывающую современные достижения молекулярной биологии и палеонтологии, автор предложил в позапрошлом году (Зеленков, 2013).
Хотя мы сейчас, несомненно, знаем о родстве между различными группами птиц намного больше, чем раньше, неясного осталось еше предостаточно. Например, не до конца понятно, какое положение на филогенетическом древе птиц занимают голуби, совы, дрофы и др. В настоящее время считается, что ответить на ряд вопросов поможет исследование всего генома (до сих пор исследовались только отдельные гены либо относительно небольшие «порции» генома).
В связи с этим так важны результаты геномного исследования филогенетического дерева птиц, опубликованные в одном из недавних выпусков Science (Jarvis et al., 2014). Авторы этой работы – очень большой коллектив ученых, исследовали полный геном у представителей всех отрядов птиц и на этом основании построили новое филогенетическое древо, которое можно назвать очередным существенным шагом вперед к пониманию родства между современными пернатыми. Нужно отметить, что полученное авторами филогенетическое дерево предоставило очень мало сюрпризов: большинство обнаруженных ими ветвей так или иначе уже были выявлены в последние десятилетия (например, те же поганки и фламинго).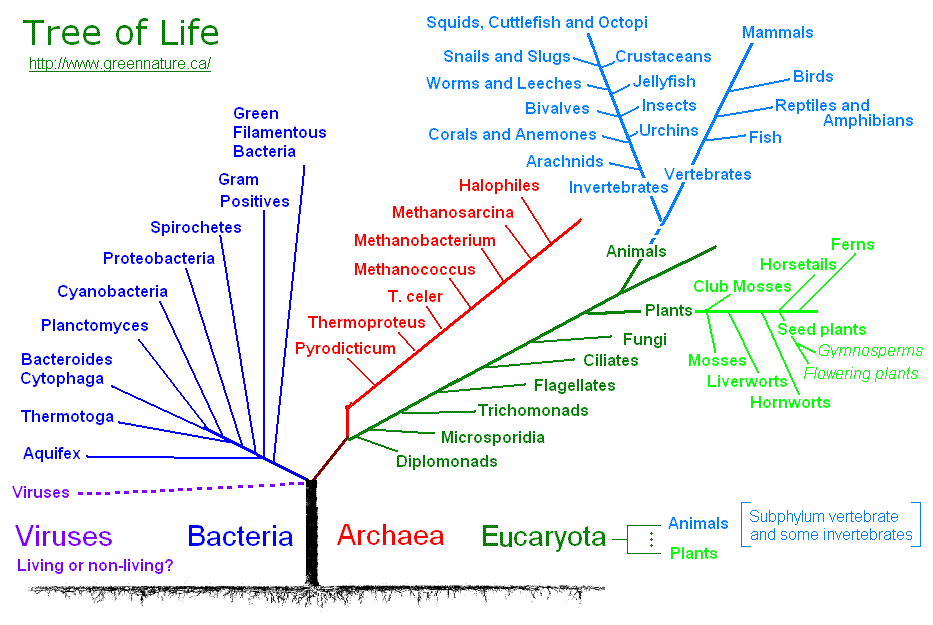 Но есть и новые интересные результаты: например, ближайшими родственниками фламинго и поганок в нашей фауне могут оказаться голуби, а ближайшими родственниками дроф – кукушки.
Но есть и новые интересные результаты: например, ближайшими родственниками фламинго и поганок в нашей фауне могут оказаться голуби, а ближайшими родственниками дроф – кукушки.
Особенно важно то, что теперь мы знаем, что современные представления о родстве между птицами базируются на основе сходства не только между отдельными генами и их комбинациями, но и всего генома. Дальнейшее развитие этого направления видится в изучении все большего числа геномов от птиц разных видов, ведь именно от этого во многом будет зависеть форма ветвления полученного филогенетического древа. Можно утверждать, что несмотря на долгий прогресс в науке об эволюции птиц, мы все еще стоим на пороге больших открытий.
Литература
Зеленков Н. В. Система птиц (Aves: Neornithes) в начале XXI века //Тр. Зоологического ин-та РАН. 2013. Прил. № 2. С. 174—190.
Jarvis E. D. et al. Whole-genome analyses resolve early branches in the tree of life of modern birds. //Science. 2014. V. 346. N. 6215. P. 1320—1331.
//Science. 2014. V. 346. N. 6215. P. 1320—1331.
Sibley C. G., Ahlquist J. E. Phylogeny and classification of birds: a study in molecular evolution. Yale University Press, New Heaven, London, 1990. 976 p.
: 30 Мар 2015 , Наша Арктика , том 61,
№1
Учные построили филогенетическое древо кошек
Устав работать с человеком, генетики взялись за кошек. Построенное ими «генеалогическое древо» показывает, что некоторые совсем не похожие друг на друга породы состоят в близком родстве, а расселение этих домашних животных по миру шло весьма причудливыми маршрутами.
Считается, что территории «Плодородного полумесяца» Ближнего Востока — земли Ирака, юго-восточной части Турции, Сирии и Палестины, а также долина нижнего течения Нила — являются колыбелью человеческой цивилизации. Считается, что именно здесь произошло и одомашнивание кошек, которое часто связывают с той особой мистической ролью, что отводилась этим животным в представлениях египтян о мире.
Генетическое исследование, проведенное специалистами из США, Израиля, Италии, Турции и Финляндии, подтвердило, что именно этот регион является родиной всех домашних кошек, а также выявило неожиданное родство между, казалось бы, мало связанными друг с другом кошачьими породами. Работа учёных опубликована в последнем номере журнала Genomics.
Результаты проведенных ранее археологических исследований, а также данные об эволюции котов позволяют предположить, что их одомашнивание произошло примерно 5–10 тысяч лет назад в областях, ныне объединенных под названием «Плодородного полумесяца» Ближнего Востока. «Плодородный полумесяц», окаймляющий с севера внутриаравийские пустыни Саудовской Аравии, Сирии и Ирака, считается родиной земледелия и скотоводства, появившихся в неолите.
close
100%
Плодородный полумесяц
условное название региона на Ближнем Востоке, в котором в зимние месяцы наблюдается повышенное количество осадков. Введено в научный оборот американским археологом и историком начала XX века Джеймсом Брэстедом.
По оценкам современных учёных, одомашнивание котов произошло тогда, когда первые люди начали переходить от кочевого образа жизни к оседлому, сопряженному с ведением сельского хозяйства и содержанием домашнего скота. Кошки, умевшие ловить мелких грызунов, оказались чрезвычайно полезными для людей, которые выращивали зерновые культуры. Кроме того, немалую роль, видимо, сыграл статус священного животного, которым кошек наделили египтяне.
В конечном итоге кошки и коты стали домашними любимцами, но в полной мере не одомашнились.
Даже сейчас весьма домашние коты проявляют самостоятельность и не прочь поохотиться — даже в том случае, если хозяева ежедневно кормят их.
С развитием человеческой цивилизации, в частности, с появлением длинных торговых маршрутов, коты начали распространяться по свету. В отличие от других домашних животных, кошачьи редко становились мишенью селекционеров, желавших получить породы, особи которых, например, будут эффективнее ловить мышей. Любителей кошек интересовал скорее вид их любимца, его эстетические характеристики, в связи с чем люди часто пытались вывести породы, отличающиеся особым цветом меха.
Из-за этого исследование кошек не так сложно с генетической точки зрения. Если для вывода собачьих пород необходимо было сложное скрещивание, подразумевающее сложные взаимодействия нескольких генов, появление новой породы кошек часто определялось выделением чистой генетической линии с сильной экспрессией того или иного гена.
Сейчас эксперты уважаемой в кошачьем мире американской Ассоциации любителей кошек (CFA) насчитывают около 50 кошачьих пород, 16 из которых признаны «естественными», то есть развившимися в определенных регионах. Остальные появились в последние полвека благодаря усилиям селекционеров.
Как отмечают руководители исследования Лесли Лайонс и Моника Липински из Университета Калифорнии в Дэвисе, свое внимание они сфокусировали на нескольких проблемах. Во-первых, их интересовало то, каким образом кошачьи путешествовали по древнему миру и попадали в такие, казалось бы, отдаленные от «Плодородного полумесяца» районы, как Северная и Южная Америки. Кроме того, учёные изучали разнообразие генетики котов, которая формировалась по мере их расселения по свету. Наконец, исследователи пытались обнаружить некоторую редукцию генетического разнообразия, которая могла произойти у представителей древних или современных пород.
Наконец, исследователи пытались обнаружить некоторую редукцию генетического разнообразия, которая могла произойти у представителей древних или современных пород.
Для проведения подобного масштабного исследования авторы собрали образцы мазков слюны за щекой более чем 1100 котов. Часть котов-доноров принадлежала 17 популяциям «дворовых котов», естественным образом размножающихся на территории Европы, Средиземноморья, Африки, Азии и обеих Америк, а часть принадлежала 22 общепризнанным породам. Среди них оказались представители и «естественных», и выведенных в последние полвека пород.
Образцы ДНК были получены либо на кошачьих конкурсах, либо высылались непосредственно хозяевами. Кроме того, учёные использовали микросателлиты (области повторов в хромосомной ДНК), чтобы определить степень генетического родства тех или иных пород, их географическое происхождение, а также степень генетических потерь, которые стали результатом инбридинга — скрещивания близкородственных животных внутри популяций.
Проведя анализ образцов ДНК, учёные пришли к выводу, что по характеристикам наследственной информации всех современных котов можно разделить на 4 группы.
Они соответствуют определенным регионам Европы, Средиземноморья, Восточной Африки и Азии.
Впрочем, между представителями этих групп существует генетическое родство. Так, американские «дворовые» коты оказались родственны «дворняжкам» Западной Европы, а короткошерстная американская кошка и кошка породы мэйн-кун (обе породы были выведены в США) являются родственными 7 другим западноевропейским породам. Это, впрочем, неудивительно, поясняют исследователи, кошки, завезенные в Новый Свет людьми, за достаточно небольшой временной отрезок не претерпели серьезных генетических изменений, которые бы привели к формированию серьезных отличий от западноевропейских кошек и котов.
Вместе с тем, западноевропейские кошки сформировали несколько плотных групп, хорошо представленных лишь в нескольких регионах. А вот Азиатская группа оказалась достаточно изолированной. По словам учёных, коты, ныне населяющие Юго-Восточную Азию и её окрестности, удалились от «Плодородного полумесяца» достаточно давно и развивались без интенсивного скрещивания с котами других пород.
По словам учёных, коты, ныне населяющие Юго-Восточную Азию и её окрестности, удалились от «Плодородного полумесяца» достаточно давно и развивались без интенсивного скрещивания с котами других пород.
Филогенетическое древо кошек, построенное на основе генетических данных // Genomics
Исследователям удалось узнать много нового и о конкретных породах кошек. Например, персидская кошка — возможно, самая древняя чистая порода — не приходится близкой родственницей котам с географически близкого ей Ближнего Востока. Как ни парадоксально, но персы генетически более близки кошачьим, проживающим на Западе Европы. Впрочем, известно, что персидская кошка в разное время и в разных странах называлась не только персидской, но и китайской, индийской и даже русской — по названиям стран, откуда они были завезены в Европу, однако в их неевропейском происхождении никто не сомневался. Возможно, в жилах современных европейских котов, несмотря на слабое внешнее сходство, немало крови «персидских» предков.
Стоит заметить также, что многие известные породы, например, персы, используются для выведения новых пород, часто более короткоголовых (имеющих короткий и широкий череп). Например, экзотические короткошерстные коты, которые, по сути, являются персами с «короткой стрижкой», представляют собой вариант новой породы, связавшей две другие, ранее отдельные группы кошек.
Кошки из Сингапура оказались помесью азиатских и западноевропейских котов, коты-итальянцы — помесью средиземноморских и европейских котов, породы из юго-восточной Азии породнились с вьетнамскими «крысоловами». А японские бесхвостые кошки, бобтейлы, также оказались родственны котам Западной Европы, хотя некоторая генетическая связь с восточной, азиатской группой у представителей породы осталась.
Измерение генетического разнообразия
Один из способов количественного измерения степени генетического разнообразия – подсчт показателя гетерозиготности. Свойством гетерозиготности называют наличие разных аллелей гена в парных хромосомах диплоидного организма.
Анализ молекулярной дисперсии позволил учёным вычислить степень генетического разнообразия западноевропейских (и американских), восточноафриканских, средиземноморских и восточноазиатских котов. Самыми разнообразными оказались средиземноморские коты (около 500 аллелей во всех маркерах), и гены именно этих котов оказались лучше всего перемешанными внутри популяции. Возможно, причиной тому служит постоянное движение караванов и судов, происходившее во время интенсивного одомашнивания и расселения котов.
А вот среди чистых пород самой генетически разнообразной оказалась сибирская кошка (показатели гетерозиготности — 0,69 и 0,73).
У неё также ниже всего показатели, численно выражающие степень инбридинга.
«Результаты нашего исследования подтверждают итоги других работ, согласно которым приручение первых кошек произошло в землях «Плодородного полумесяца». Кроме того, наше исследование полезно для людей, разводящих новые породы», — рассказала первый автор исследования, аспирантка Моника Липински.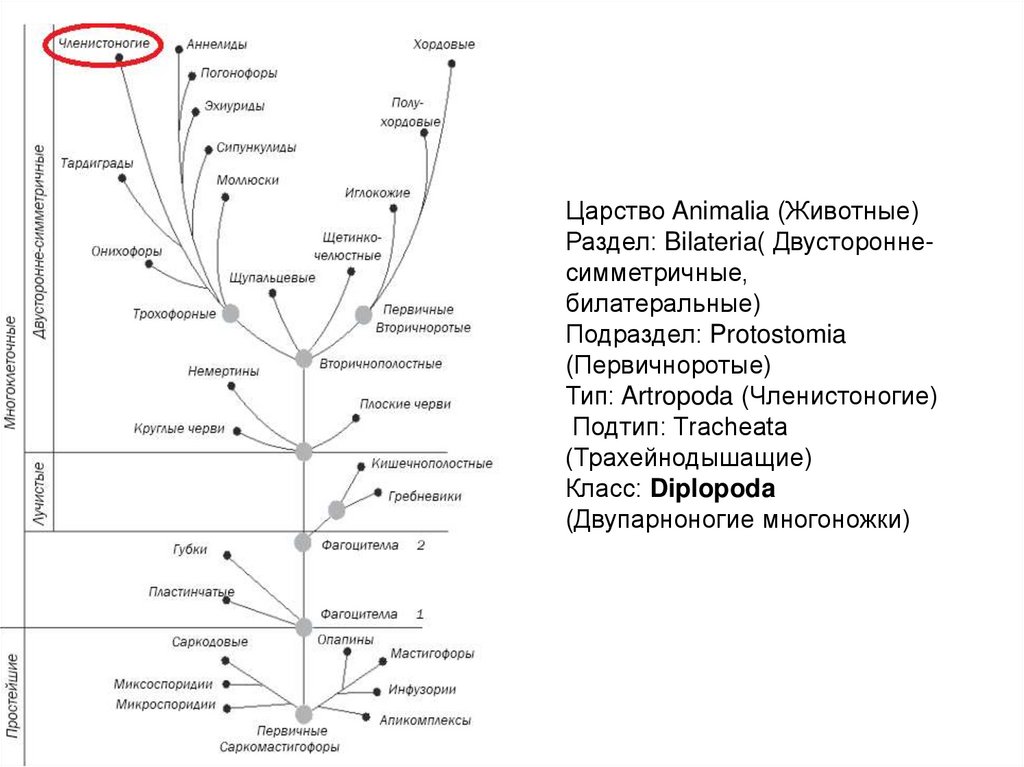
По словам профессора Лесли Лайонс, курировавшей исследование, в ходе работы учёные обнаружили более 200 генетических нарушений, многие из которых были зафиксированы у представителей чистых пород. Подобная информация, по мнению Лайонс, должна дойти до людей, занимающихся разведением котов, дабы они не создавали породы заведомо больных кошек.
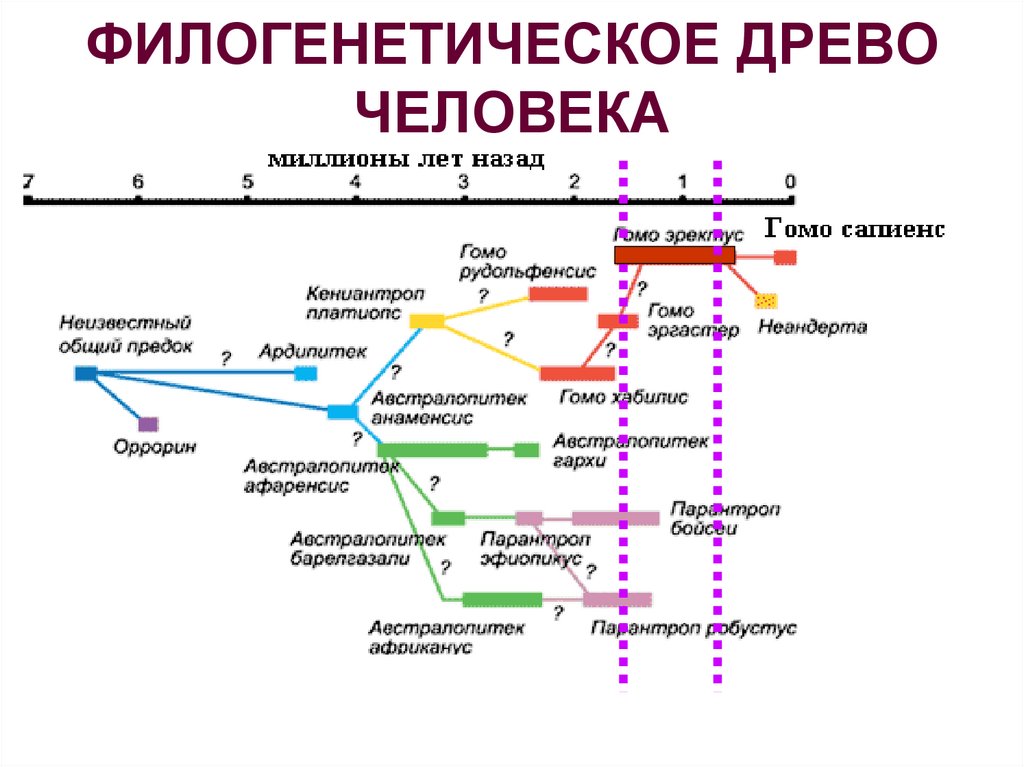
44.Современные взгляды на филогенетическое древо человека.
Систематика
– это наука о биологической классификации.
Она включает изучение органического
разнообразия и даёт средства для изучения
исторических аспектов эволюции.
В
современной систематике традиционно
выделяют три направления – эволюционную
систематику, фенетику и филогенетическую
систематику (кладистику)
Эволюционная
систематика
Согласно
этой систематике изучение популяций и
внутривидовой изменчивости имеет
решающее значение для понимания
биологического разнообразия. Этот
подход изложен в работах орнитолога
Эрнста Майра.
Палеонтологический
взгляд на биологическую систематику,
тесно связанный с «эволюционной
систематикой», изложил Джордж Гейлорд
Симпсон . Симпсон, будучи специалистом
Симпсон, будучи специалистом
по ископаемым млекопитающим, уделял
много внимания взаимосвязям между
группами организмов.Он полагал, что
конструирование классификаций – это
полезный результат таксономических
исследований.Цель таксономии понять,
как взаимосвязаны и группируются
организмы с биологической точки зрения.
Симпсон считал, что таксономия является
наукой.
Классическая
эволюционная систематика стремится
внести максимальное количество
эволюционной информации в биологические
классификации. Формальная классификация
является попыткой в максимальной степени
и одновременно отразить генетическое
сходство и филогению, причем полное
соответствие между классификацией и
филогенией невозможно.
Фенетика
В
конце 1959-х появились работы, в которых
классификация основывалась на концепции
общего сходства. Главными приверженцами
этого подхода были Сокел и Снит. Хотя
это направление в систематике было
сначала названо «нумерической
таксономией», термин «фенетика»,
предложенный Майром в 1965 году более
предпочтителен, т.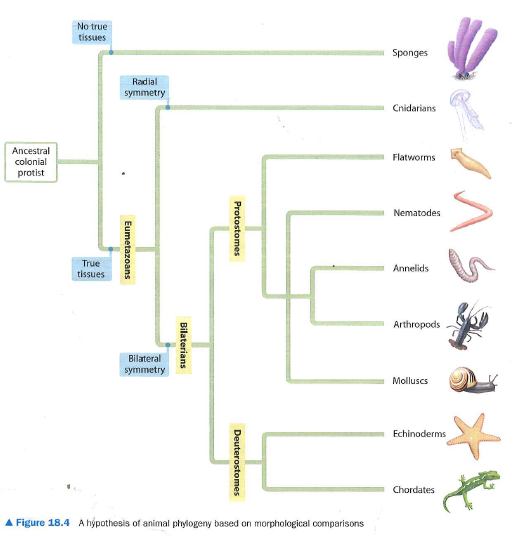 к количественные
к количественные
подходы в таксономии существуют и в
других направлениях систематики.
Сторонники фенетики стремились сделать
таксономию объективной, причём главная
её цель – создавать классификации, в
которых связи между группами организмов
определяются на основе общего сходства.
Эволюционное содержание полностью
исключается из таксономии, так как в
большинстве случаев филогения
неизвестна.
Фенетический
подход основывается на том, что следует
использовать максимальное число
невзвешенных (то есть считающихся
равноценными) наблюдений для построения
классификации, что делает её независимой
от какой-либо теории, например от теории
органической эволюции. Тогда процесс
сбора данных становится непредвзятым
и может быть автоматизирован с помощью
компьютеров.
Фенетические
методы заключаются в преобразовании
числа различий в «расстояния», которые
служат способом измерения сходства
между группами организмов. Эти расстояния
вычисляются путём подсчета числа
различий признаков между всеми возможными
парами таксонов.
Филогенетика
(кладистика)
Немецкий
энтомолог и теоретик систематики Вилли
Хенниг предложил ряд методов и принципов
в своей работе.
Хенниг
предложил подход, который отражает
информацию о результатах эволюционного
процесса. Хенниг рассматривал иерархические
классификации как общую справочную
систему биологии. Вместе с тем, он
утверждал, что полезность этих
классификаций может быть максимальной
только в том случае, если они максимально
отражают филогенетические отношения
этих организмов. Филогенетические
отношения рассматриваются исключительно
как генеалогические без учета степени
дивергенции. Эта точка зрения не была
принята ни фенетиками, ни эволюционными
систематиками. Хеннигов подход,
обозначенный сначала как «филогенетичсеская
систематика», а теперь называемый
«кладистикой», ясно выразил идею, что
генеалогические отношения должны
основываться на уникальном сходстве
(уникальных признаках). Эти отношения
должны быть точно отражены в формальной
иерархической классификации. Все
Все
ли признаки организмов полезны для
построения классификации? Фенетики
полагают, что ответ на этот вопрос должен
быть положительным. Они используют
методы, которые оценивают степень
сходства и различия между группами.
Кладисты придерживаются точки зрения,
что группы могут формироваться только
на основе уникальных признаков. Многие
эволюционные систематики принимают
хеннигову кладистическую точку зрения
о возникновении уникальных признаков
в определенных группах в ходе эволюции,
но считают, что при установлении ранга
в формальной классификации должна
учитываться степень различия между
группами по многим признакам.
|
Филогенетические деревья — объяснение с примерами
Знания
Знайте все о филогенетических деревьях
Определение филогенетического дерева
Филогенетическое дерево (обычно известное как эволюционное дерево или филогения ) представляет собой диаграмму с ветвями, которая отображает эволюционные отношения между различными биологическими видами.
Источник: khanacademy.org
История филогенетических деревьев
Поскольку филогенетическое древо известно как « древо жизни », это усовершенствованное понимание старой концепции, которая гласит, что жизнь продвигается от более низких к более высоким уровням, как лестница, ссылаясь на то, что это похоже на Великую Цепь Бытия. . Ранние представления, наброски и наброски представляют собой ветвящееся филогенетическое древо, составляющее палеонтологическую схему. Это прежде всего указывало на геологические ассоциации и корреляции между растениями и животными.
Части филогенетического дерева
Простое филогенетическое дерево состоит из нескольких частей, упомянутых ниже:
Таксоны
Таксоны известны как «кончики» ветвей деревьев или проводимого исследования. Вы можете найти «Таксоны» на любом систематическом уровне с указанием видов, отрядов или популяций. В дополнение к этому, этот «Таксон» называется «OTU», что сокращенно означает «Оперативные таксономические единицы».
В дополнение к этому, этот «Таксон» называется «OTU», что сокращенно означает «Оперативные таксономические единицы».
ветви
Систематические и/или продолжающиеся линии дерева известны как его ветви, представляющие взаимосвязь между одним или несколькими объектами.
Узел
Кончики этих ветвей или точки, где все эти ветви соединяются вместе, известны как «узлы». Внутри узлов есть два типа: «Внутренние» и «Внешние» узлы. Предполагается, что внутренние узлы соединяют ветви, тогда как внешние узлы представляют собой углы, соединяющие таксоны вместе.
Рут и Клейд
Кроме того, у некоторых деревьев есть базальный узел, известный как «корень» дерева. Затем идет «Клада», широко известная как «предок» всех ответвляющихся видов.
Сильные стороны и ограничения филогенетического дерева
Как и любая систематическая модель, парадигма филогенетического дерева имеет некоторые сильные и слабые стороны, которые упомянуты ниже.
Сильные стороны
- Модель демонстрирует историческое происхождение любой единицы, индивидуума или вида.
- Филогенетическое древо не ранжирует виды, в отличие от линнеевской модели, которая искусственно ранжирует организмы по царствам. Известно, что это искажение, потому что оно показывает, что разные группы с общими рангами эквивалентны.
- Филогенетические деревья, как правило, дают представление об исторических событиях различных организмов.
- Эта модель помогает в изучении развития и прогрессии всех существующих юнитов.
Ограничения
- Филогенетическое древо демонстрирует гипотезы развития и развития жизни.
- Они точны ровно настолько, насколько точны данные, на которых они основаны и продемонстрированы.
- Данные получены из исследований и исследований, которые в некоторой степени предвзяты.
- Следовательно, филогенетические деревья, основанные на данных исследований и исследований, всегда могут быть необъективными, манипулируемыми или неточными.

- Это прогрессивное лестничное филогенетическое древо черепов развилось из организма, не имевшего позвоночного столба.
- Следовательно, организмы, объекты или единицы с разными характеристиками помещаются в разные группы на каждом конце ветви исключительно на основе общих характеристик.
Типы филогенетических деревьев
Дендрограмма
Дерево обычно известно как Дендрограмма. Это не обязательно филогенетическое дерево, оно используется для представления и демонстрации чего-либо схематически.
Хронограмма
Хронограмма — это тип филогенетического дерева, которое представляет время через длину своих ветвей.
Филограмма
Как следует из названия, филограмма — это филогенетическое дерево, длина ветви которого пропорциональна степени изменения признака.
кладограмма
Известно, что этот тип филогенетического дерева демонстрирует только схему ветвления.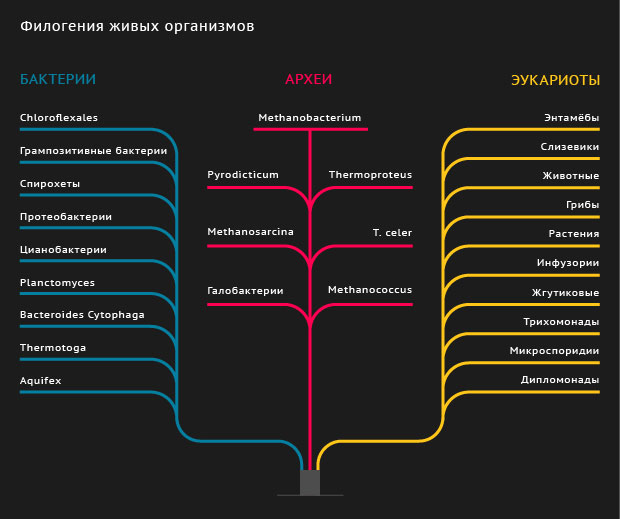 Это означает, что в его ветвях отсутствует представление о времени изменения характера. Кроме того, его внутренние узлы не представляют его предков.
Это означает, что в его ветвях отсутствует представление о времени изменения характера. Кроме того, его внутренние узлы не представляют его предков.
Дальгренограмма
Этот тип филогенетического дерева демонстрирует поперечное сечение.
Филогенетическая сеть
Филогенетическая сеть не похожа на дерево, а представляет собой общую диаграмму или граф. Он в основном используется для преодоления недостатков или ограничений, присущих деревьям.
Источник: pnas.org
Схема шпинделя
Диаграмма веретена также известна как пузырьковая диаграмма, используемая для представления таксономического разнообразия в зависимости от геологического времени. Это отражает изменение обилия нескольких таксономических категорий во времени.
Различия между филогенезом, эволюционным деревом, филогенетическим деревом и кладограммой
| Филогенез | Эволюционное дерево | Филогенетическое дерево | Кладограмма |
|---|---|---|---|
Филогенез — это раздел биологии, специально занимающийся филогенезом. | Эволюционное дерево — это наглядная демонстрация эволюции видов от точки их происхождения. | Филогенетическое дерево представляет собой схематическое изображение развития биологических видов. | Это разветвленное представление, которое изображает кладистические отношения между всеми видами видов. |
Как читать филогенетическое дерево?
Чтение филогенетического дерева аналогично чтению некоторых генеалогических деревьев. Тем не менее, мы дадим вам знать, как читать филогенетическое дерево шаг за шагом.
Шаг 01. Увидьте корень как предка, а подсказки как потомков
Корень дерева означает общего предка, от которого произошли все виды. А кончик означает группу видов (часть общего предка).
Шаг 02: См. родословную дочерей
Второй шаг – определить дочерние линии, происходящие от общего предка. Он разделяется на две или более ветвей, создавая линию различия между одним или несколькими видами.
Шаг 03: См. общую родословную между родословными (если есть)
Третий шаг — выяснить, существуют ли какие-либо общие родословные между группой видов. См. точки B и C на рисунке ниже. Точки B и C имеют уникальную историю, в отличие от точки A.
Шаг 04: Прочтите диаграмму в целом
Посмотрите на рисунок ниже, и теперь вы сможете прочитать его целиком. Оранжевый цвет показывает, что у точек A, B и C есть общий предок. Однако зеленый цвет показывает, что у B и C один общий предок, и то же самое касается точки C.
Как составить филогенетическое дерево?
Вы можете построить филогенетическое дерево, выполнив следующие шаги.
Шаг 01: Определите виды предков
Первый шаг заключается в выявлении самых разных видов. Обратите внимание, что он будет иметь наибольшее количество мутационных отличий от других видов с наибольшим числом.
Шаг 02: Определите следующий самый отличающийся вид
Следующим шагом будет поиск следующих самых разных видов, имеющих общего предка с предыдущими видами. Назовем его А.
Назовем его А.
Шаг 03: начните рисовать филогенетическое дерево
Начните рисовать общую линию и продлите ее ветви, чтобы показать возможного общего предка A ветвей B и C.
Шаг 04: Начните добавлять организмы
Найдите характеристики другого организма и найдите организм, наиболее характерно похожий на этот конкретный ген.
Шаг 05: Добавьте оставшиеся оргазмы
Следуйте тому же методу и начните добавлять оставшиеся организмы, просматривая строки и столбцы и находя генетически идентичные организмы.
Примеры для филогенетического древа жизни
Для вашего понимания, здесь у нас есть примеры филогенетических деревьев человека и филогенетических деревьев животных.
Филогенетическое древо человека
Как вы можете видеть на диаграмме, у каждого вида или особи (в данном случае) есть общий предок, и это ваш дедушка и бабушка. Затем он разделяется на две ветви: ваш родитель и ваша тетя (брат или сестра вашего родителя).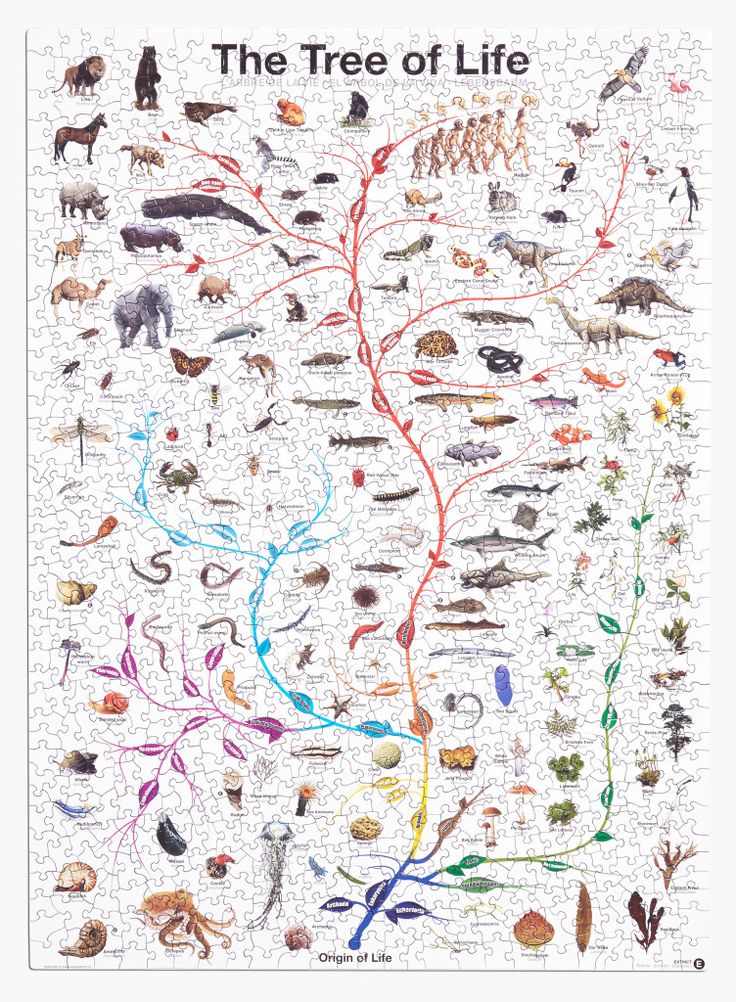 И тогда у вас, вашего брата, сестры и двоюродных братьев есть уникальная история, потому что вы родились от разных родителей, но у вас один и тот же общий предок, что и у вашего дедушки и бабушки.
И тогда у вас, вашего брата, сестры и двоюродных братьев есть уникальная история, потому что вы родились от разных родителей, но у вас один и тот же общий предок, что и у вашего дедушки и бабушки.
Филогенетическое дерево животных
Как видно на схеме, у всех животных один общий предок, но они разделены из-за разных характеристик. Эти характеристики — челюсти, легкие, желудки и перья. Таким образом, эти характеристики различают животных, упомянутых на диаграмме, которые, тем не менее, имеют одного и того же предка.
Завершение статьи!
В целом, филогенетическое дерево можно использовать и для отображения отношений между другими объектами, которые в первую очередь основаны на сходствах и различиях их характеристик и указывают на то, что все виды, объекты, индивидуумы и единицы укоренены. в общем происхождении. Дерево может быть нарисовано по-разному, и это может быть виртуальное или даже физическое.
Однако для виртуального моделирования диаграммы филогенетического дерева лучше всего подходит удобное программное обеспечение EdrawMax , которое помогает создавать 2D-диаграммы, блок-схемы, диаграммы Ганта и все виды филогенетических деревьев и моделей.
Связанные статьи
Программное обеспечение для архитектурного макета
Матрица Ансоффа
Диаграмма анатомии человека
Все о модном дизайне
Что такое алгоритм
Административно-организационная структура Определение
2.1 Чтение деревьев — цифровой атлас древней жизни
Части дерева
Филогенетическое дерево — это иллюстрация, изображающая гипотетические степеней эволюционных взаимоотношений между выбранным набором таксонов (единственное число = таксон). Таксоны, как правило, являются видами, но также могут быть линнеевскими группами более высокого уровня, такими как роды или семейства. В качестве альтернативы, некоторые филогенетические деревья изображают отношения между особями внутри вида (например, из географически изолированных популяций). Независимо от их ранга, таксоны, изображенные на филогенетическом дереве, часто называют терминальными таксонами, потому что они встречаются на концах дерева. Их иногда называют «терминалами» или «листьями».
Таксоны, как правило, являются видами, но также могут быть линнеевскими группами более высокого уровня, такими как роды или семейства. В качестве альтернативы, некоторые филогенетические деревья изображают отношения между особями внутри вида (например, из географически изолированных популяций). Независимо от их ранга, таксоны, изображенные на филогенетическом дереве, часто называют терминальными таксонами, потому что они встречаются на концах дерева. Их иногда называют «терминалами» или «листьями».
Терминальные таксоны связаны ветвями. Ветви — это отрезки линий, из которых состоит дерево. Ветви сходятся в точках ветвления, называемых узлами. Каждый узел представляет общего предка, разделяемого двумя или более конечными таксонами.
Части филогенетического дерева, включая конечные таксоны, ветви и узлы. Изображение Джонатана Р. Хендрикса (Creative Commons Attribution-Sharealike 4.0 International license).
Родство таксонов
Помните, что филогенетические деревья изображают градусов родства между таксонами.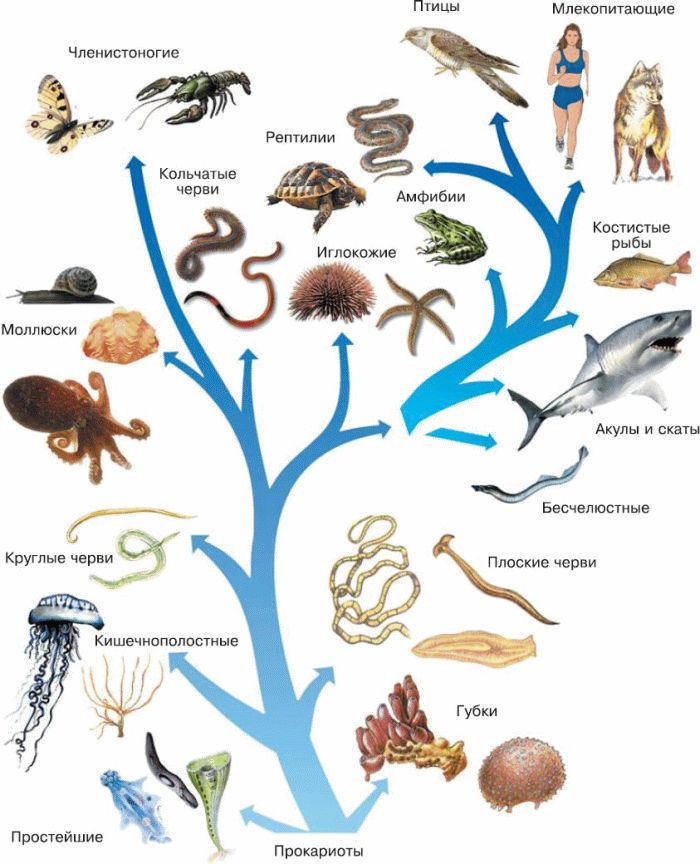 На филогенетическом дереве более близкородственные терминальные таксоны связаны более мелкими узлами (т. е. узлами ближе к вершинам дерева), а более отдаленно связанные терминальные таксоны связаны более глубокими узлами (т. е. узлами ближе к основанию дерева). .
На филогенетическом дереве более близкородственные терминальные таксоны связаны более мелкими узлами (т. е. узлами ближе к вершинам дерева), а более отдаленно связанные терминальные таксоны связаны более глубокими узлами (т. е. узлами ближе к основанию дерева). .
Посмотрите на рисунок выше. На этом рисунке таксон B и таксон C более тесно связаны друг с другом, чем с таксоном A. Мы знаем это, потому что таксон B и таксон C имеют более мелкий узел (синий узел), чем узел, который является общим с таксоном A. (желтый узел). Другой способ изложить это наблюдение состоит в том, что таксон B и таксон C имеют общий узел (синий узел), который не является общим с таксоном A. Взяв более широкий взгляд, мы также знаем, что таксоны A, B и C более тесно связаны друг с другом. кроме таксонов D, E, F, G и H. Это связано с тем, что таксоны A, B и C имеют общий желтый узел, который не связан с таксонами D, E, F, G или H. . Таксоны A, B и C связаны с таксонами D, E, F, G и H более глубоким узлом (красным узлом).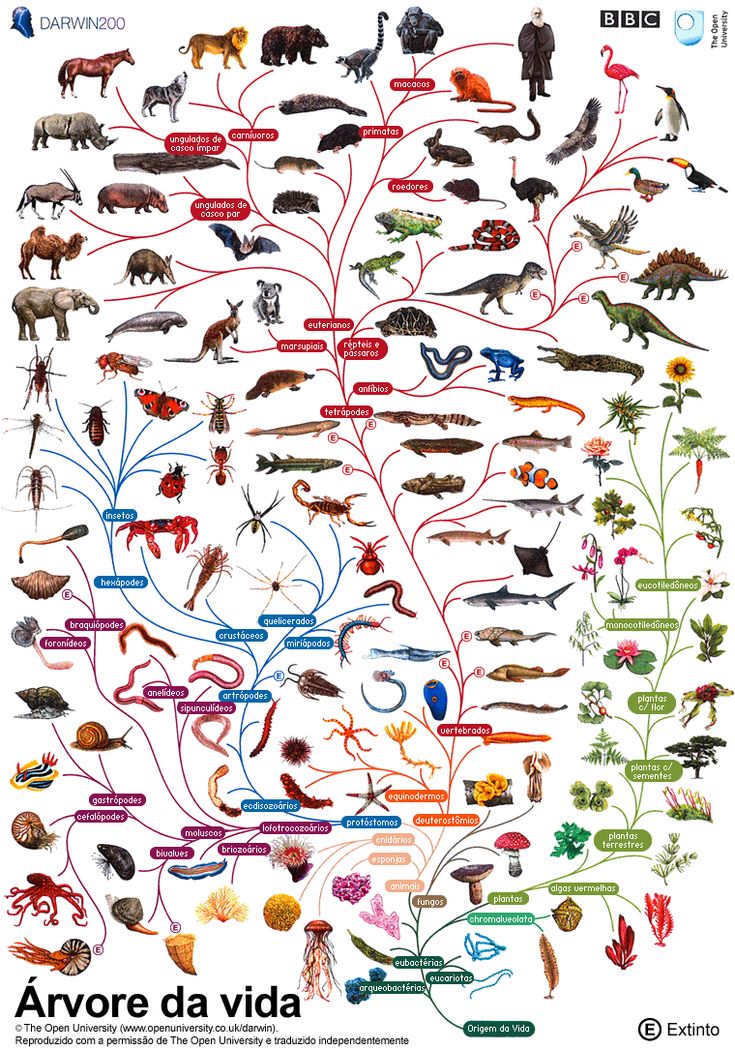
Ветви могут вращаться вокруг узлов без каких-либо изменений в гипотетических отношениях, изображенных на древовидной диаграмме. Убедите себя, что три приведенных ниже дерева изображают точно такой же набор взаимоотношений между таксонами A-H. В каждом случае единственное изменение, которое было сделано, состоит в том, что ветви и связанные с ними конечные таксоны были повернуты вокруг узлов. Помните, что степень родства между различными комбинациями терминальных таксонов определяется относительной глубиной узлов, которые их соединяют.
Дерево A. Изображение Джонатана Р. Хендрикса (Creative Commons Attribution-Sharealike 4.0 International License).
Дерево B. Изображение Джонатана Р. Хендрикса (Creative Commons Attribution-Sharealike 4.0 International License).
Дерево C. Изображение Джонатана Р. Хендрикса (Creative Commons Attribution-Sharealike 4.0 International License).
Графический стиль филогенетических деревьев варьируется. Например, дерево, показанное ниже, изображает точно такой же образец взаимоотношений между таксонами A–H, что и три дерева, показанные выше.
Например, дерево, показанное ниже, изображает точно такой же образец взаимоотношений между таксонами A–H, что и три дерева, показанные выше.
Филогенетическое дерево в виде скобок. Изображение Джонатана Р. Хендрикса (Creative Commons Attribution-Sharealike 4.0 International License).
Все чаще можно увидеть филогенетические деревья в форме круга, такие как показанное ниже. Круглые деревья часто используются для иллюстрации отношений между членами основных групп существующих организмов, и эти деревья могут иметь много терминальных таксонов. Круговые деревья можно читать так же, как деревья, показанные выше, потому что относительная глубина узлов указывает на степень родства между терминалами.
Филогенетическое древо в форме круга, на котором показаны отношения между основными группами живых организмов (синий = бактерии, зеленый = археи, розовый = эукариоты). Это дерево было создано Ивицей Летунич и воссоздано Марианой Руис Вильярреал (общественное достояние).
Клады и сестринские группы
Клада (от греческого кладос = ветвь) — это группа, включающая предка (узел) и всех его потомков (все более мелкие узлы и конечные таксоны, происходящие от этого узла) на филогенетическое дерево. Если вы выберете узел на филогенетическом дереве, вы можете легко нарисовать круг вокруг клады, которую он определяет, как в дереве ниже. Хотя многие клады не имеют формальных названий, некоторые важные клады названы в формальных схемах классификации (подробнее о кладах и классификации см. В этом следующем разделе).
Филогенетическое дерево, иллюстрирующее концепцию клад. Обратите внимание, что клады не исключают друг друга, а вложены друг в друга. Изображение Джонатана Р. Хендрикса (Creative Commons Attribution-Sharealike 4.0 International license).
Клады не исключают друг друга, а образуют вложенные наборы на дереве. Таким образом, любой данный таксон может принадлежать многим кладам. Например, в приведенном выше дереве таксон B принадлежит трем кладам: кладе, определяемой узлом 1, более инклюзивной кладе, определяемой узлом 2, и еще более инклюзивной кладе, определяемой узлом 7. Таксон E принадлежит четырем кладам, определяемым узлом 7. Узлы 3, 5, 6 и 7 соответственно. Попробуйте выяснить, к скольким кладам принадлежат таксон A и таксон H, и определить, какие узлы определяют каждую кладу.
Таксон E принадлежит четырем кладам, определяемым узлом 7. Узлы 3, 5, 6 и 7 соответственно. Попробуйте выяснить, к скольким кладам принадлежат таксон A и таксон H, и определить, какие узлы определяют каждую кладу.
Сестринские таксоны или сестринские группы представляют собой пары терминальных таксонов и/или клад, которые ответвляются от общего узла и часто считаются близкородственными. Пары сестринских терминальных таксонов на рисунке выше включают: B и C, E и F, а также G и H. Клада, определяемая узлом 3 (узел 3 + таксон E + таксон F), является сестрой клады, определяемой узлом 4 ( узел 4 + таксон G + таксон H). Терминальный таксон A является сестрой клады, определяемой узлом 1 (узел 1 + таксон B + таксон C). Попробуйте найти больше сестринских пар на дереве выше.
Значение длин ветвей
На ветвях филогенетического дерева имплицитно происходят две вещи. Во-первых, это течение времени. Более глубокие узлы старше, чем более мелкие узлы, к которым они подключены.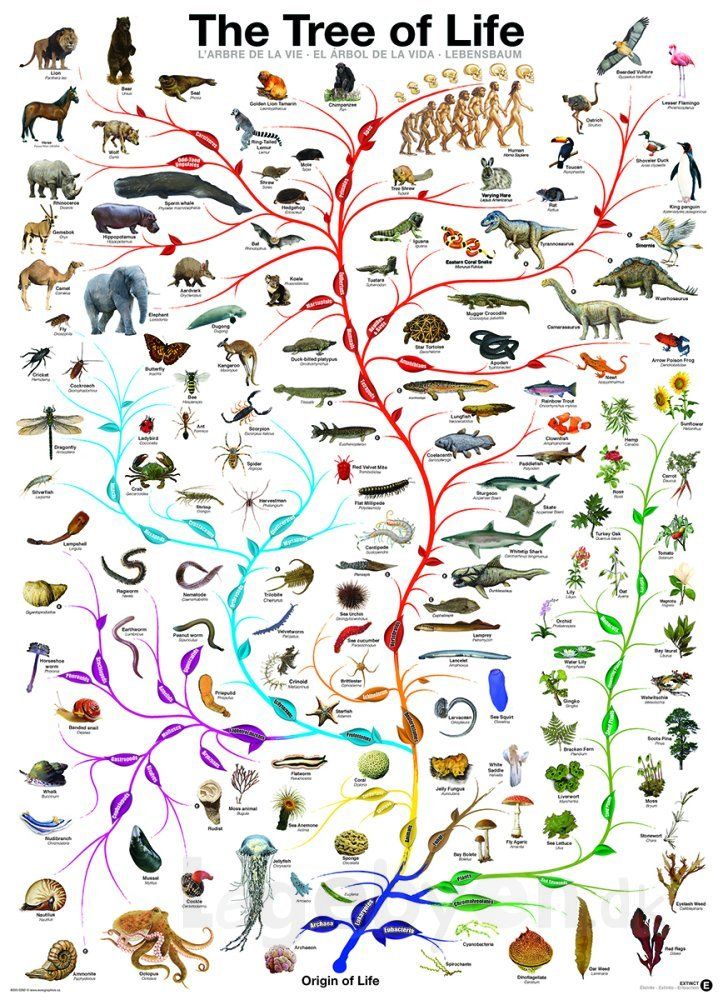 Таким образом, более глубокие узлы указывают как на более отдаленные отношения между конечными таксонами, которые они соединяют, так и на больший возраст самого последнего общего предка этих таксонов. Второе — это эволюционная модификация, или накопление наследственных генетических и/или структурных изменений по ветвям. Хотя эти изменения часто не отображаются (наносятся на карту) непосредственно на ветвях, именно эти предполагаемые изменения лежат в основе построения и интерпретации филогенетического дерева. Когда систематики говорят о «длине ветвей», они обычно имеют в виду количество этих изменений.
Таким образом, более глубокие узлы указывают как на более отдаленные отношения между конечными таксонами, которые они соединяют, так и на больший возраст самого последнего общего предка этих таксонов. Второе — это эволюционная модификация, или накопление наследственных генетических и/или структурных изменений по ветвям. Хотя эти изменения часто не отображаются (наносятся на карту) непосредственно на ветвях, именно эти предполагаемые изменения лежат в основе построения и интерпретации филогенетического дерева. Когда систематики говорят о «длине ветвей», они обычно имеют в виду количество этих изменений.
Итак, означает ли что-нибудь длина ветвей, изображенная на филогенетическом дереве (другими словами, длина ветвей на реальной диаграмме, показывающей гипотезу об эволюционных отношениях)? Ответ: это зависит.
Время и количество эволюционных изменений могут не иметь прямого отношения к относительной длине ветвей, изображенных на дереве. Многие такие деревья представляют собой кладограммы или диаграммы ветвления, сделанные с использованием кластических методов, корни которых уходят в работы Вилли Хеннига.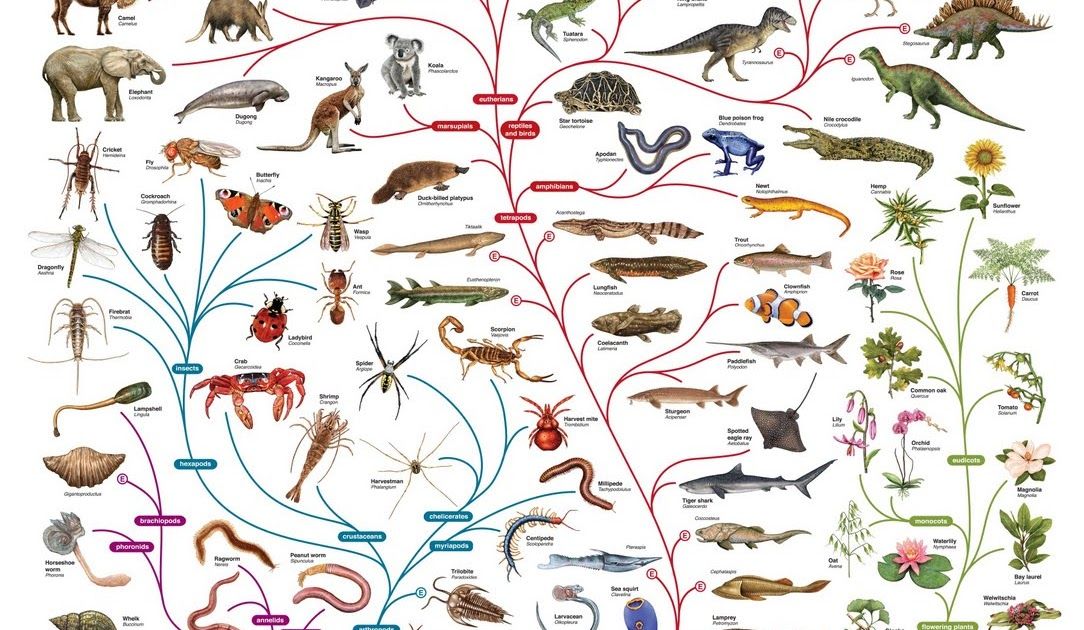 (Примечание: термин «кладограмма» иногда применяется к любому типу филогенетического дерева.) Часто диаграммы, которые рисуются для общих информационных целей, чтобы изобразить консенсусную гипотезу отношений между группой таксонов (например, в учебнике) также не имеют ветвей, масштабированных по времени или количеству эволюционных изменений. На диаграмме этого типа таксоны будут либо выровнены по концам ветвей, либо все ветви будут примерно одинаковой длины (это означает, что таксоны не выровнены).
(Примечание: термин «кладограмма» иногда применяется к любому типу филогенетического дерева.) Часто диаграммы, которые рисуются для общих информационных целей, чтобы изобразить консенсусную гипотезу отношений между группой таксонов (например, в учебнике) также не имеют ветвей, масштабированных по времени или количеству эволюционных изменений. На диаграмме этого типа таксоны будут либо выровнены по концам ветвей, либо все ветви будут примерно одинаковой длины (это означает, что таксоны не выровнены).
Филогенетическое древо, в котором ветви не привязаны ко времени или эволюционным изменениям. Эта диаграмма представляет собой кладограмму. Источник: Тернер и др. (2017) PLoS ONE 12(2): e0169885. Используется в соответствии с лицензией Creative Commons Attribution 4.0 International (CC BY 4.0).
В других случаях ветви дерева масштабируются таким образом, чтобы они отражали количество произошедших эволюционных изменений (другими словами, количество модификаций характеристик). На диаграмме этого типа длины ветвей будут различаться, и таксоны не будут выровнены по концам ветвей. Иногда этот тип дерева называют филограммой.
На диаграмме этого типа длины ветвей будут различаться, и таксоны не будут выровнены по концам ветвей. Иногда этот тип дерева называют филограммой.
Пример филогенетического дерева с ветвями, масштабированными для отображения количества эволюционных изменений (филограмма). Обратите внимание, что длины ветвей на диаграмме различаются, а таксоны не выровнены. Источник: Zapata et al. (2015) PLoS ONE 10(10): e0139068. Используется в соответствии с Creative Commons 0 1.0 Universal, Public Domain Dedication (CC0 1.0).
Деревья также могут иметь длину ветвей, масштабируемую по времени, что делает взаимосвязь между относительной глубиной узла и временем явной. Как правило, шкала времени (относительная и/или числовая) будет включена рядом с деревом, чтобы указать время событий ветвления. Если дерево явно масштабировано по времени, его можно назвать хронограммой; такие деревья также иногда называют «деревьями времени» (также временными деревьями или временными деревьями). Если все таксоны в хронограмме сохранились (живут), они будут выровнены в настоящее время.
Если все таксоны в хронограмме сохранились (живут), они будут выровнены в настоящее время.
Пример филогенетического дерева с ветвями, масштабированными для отображения времени (хронограмма). Серые полосы в узлах — это полосы ошибок. Источник: Чжан и др. (2013) PLoS ONE 8(7): e70449. Используется в соответствии с лицензией Creative Commons Attribution (CC BY).
Иногда вымершие таксоны могут быть включены в качестве конечных точек филогенетического дерева. Если такое дерево имеет ветви, масштабированные по времени, вымершие таксоны не будут выровнены в настоящее время. Вероятнее всего, кончики ветвей вымерших таксонов закончатся на уровне времени, на котором они вымерли, как показано ниже.
Примеры кладограмм, масштабированных по времени и включающих вымершие таксоны в качестве терминалов. Из: Райт и Стигал (2013) PLoS ONE 8(7): e68353. Используется в соответствии с лицензией Creative Commons Attribution (CC BY).
Обзор
Вопрос
Какой таксон на этом филогенетическом дереве является родственным таксону B?
Ответ
Таксон C.
Вопрос
Какой таксон на этом филогенетическом дереве является родственным таксону D?
Ответ
Клада, включающая таксоны E, F, G и H.
Вопрос
В этом филогенетическом дереве идентифицированный узел представляет общего предка какого таксона?
Ответ
Таксоны D, E, F, G и H.
Ссылки
Turner, A. H., A. C. Pritchard, and N. J. Matzke. 2017. Эпирический и байесовский подходы к времени расхождения только по ископаемым: исследование трех клад рептилий. PLoS ONE 12(2): e0169885. https://doi.org/10.1371/journal.pone.0169885.
Райт, Д. Ф. и А. Л. Стигал. 2013. Геологические факторы изменения фауны в позднем ордовике в Лаврентии: исследование связей между тектоникой, видообразованием и биотическими инвазиями. PLoS ONE: 8(7): e68353. https://doi.org/10.1371/journal.pone.0068353.
Zapata, F., F.E. Goetz, S.A. Smith, M. Howison, S. Siebert, S.H. Church, S.M. Sanders, C.L. Ames, C.S. McFadden, S.C. France, M. Daly, A.G. Collins, S.H.D. Haddock, C.W. Dunn и П. Картрайт. 2015. Филогеномный анализ подтверждает традиционные отношения внутри Cnidaria. ПЛОС ОДИН 10(10): e0139068. https://doi.org/10.1371/journal.pone.0139068.
France, M. Daly, A.G. Collins, S.H.D. Haddock, C.W. Dunn и П. Картрайт. 2015. Филогеномный анализ подтверждает традиционные отношения внутри Cnidaria. ПЛОС ОДИН 10(10): e0139068. https://doi.org/10.1371/journal.pone.0139068.
Чжан, Ж.-Б., Р.-К. Ли, X.-G. Сян, С.Р. Манчестер, Л. Линь, В. Ван, Дж. Вен и З.-Д. Чен. 2013. Интегрированные ископаемые и молекулярные данные показывают биогеографическую диверсификацию восточноазиатско-восточно-североамериканского дизъюнктного рода гикори ( Carya Nutt.). PLoS ONE 8(7): e70449. https://doi.org/10.1371/journal.pone.0070449.
Дополнительные ресурсы
Следующие две статьи рекомендуются для получения дополнительной информации о чтении филогенетических деревьев:
Баум Д., С. ДеВитт Смит, С. С. С. Донован. 2005. Задача древовидного мышления. Наука 310 (5750): 979-980. Также обратите внимание, что с этой статьей связана викторина, доступ к которой можно получить здесь.
Баум, Д. 2008. Чтение филогенетического дерева: значение монофилетических групп.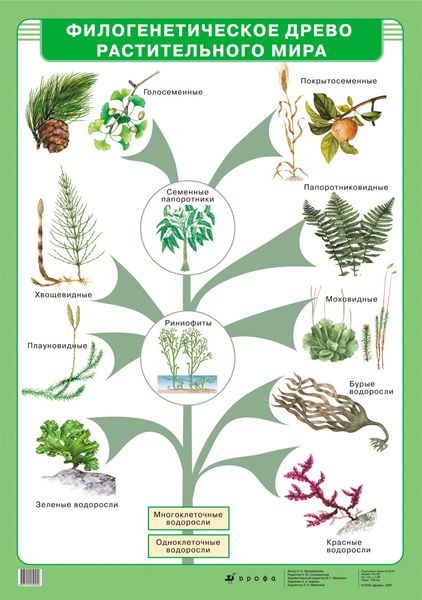 Природоведение 1(1):190.
Природоведение 1(1):190.
Использование контента
Использование текста и изображений, созданных для DEAL: Текст на этой странице написан Элизабет Дж. Хермсен и Джонатаном Р. Хендриксом. Исходный письменный контент, созданный Элизабет Дж. Хермсен и Джонатаном Р. Хендриксом для Цифровой энциклопедии древней жизни, представленный на этой странице, находится под лицензией Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. Оригинальные изображения, созданные Джонатаном Р. Хендриксом, также находятся под лицензией Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
Контент, полученный с других веб-сайтов: Атрибуция, исходная веб-страница и информация о лицензировании или условия использования указаны для изображений, полученных с других веб-сайтов, в подписи к рисунку под соответствующим изображением. Подробности смотрите в первоисточниках. Для встроенных видео указываются атрибуция и исходная веб-страница. Условия использования см. в первоисточниках. Воспроизведение изображения или видео на этой странице не подразумевает их одобрения автором, создателем, исходным веб-сайтом, издателем и/или правообладателем.
Условия использования см. в первоисточниках. Воспроизведение изображения или видео на этой странице не подразумевает их одобрения автором, создателем, исходным веб-сайтом, издателем и/или правообладателем.
FigTree
FigTree
FigTree предназначена для графического просмотра филогенетических деревьев и программы для создания готовых к публикации рисунков. Как и большинство моих программ, она была написана для моих собственных нужд, поэтому может быть не такой отточенной и многофункциональной, как коммерческая программа. В частности, он предназначен для отображения сводных и аннотированных деревьев, созданных BEAST.
Скомпилированные двоичные файлы (для Mac, Windows и Linux) доступны в репозитории FigTree GitHub.
Исходный код доступен на GitHub.
| Подпишитесь на список рассылки объявлений Figtree: |
| Электронная почта: |
| Посетите эту группу |
Этот список рассылки предназначен только для объявлений о новых версиях FigTree.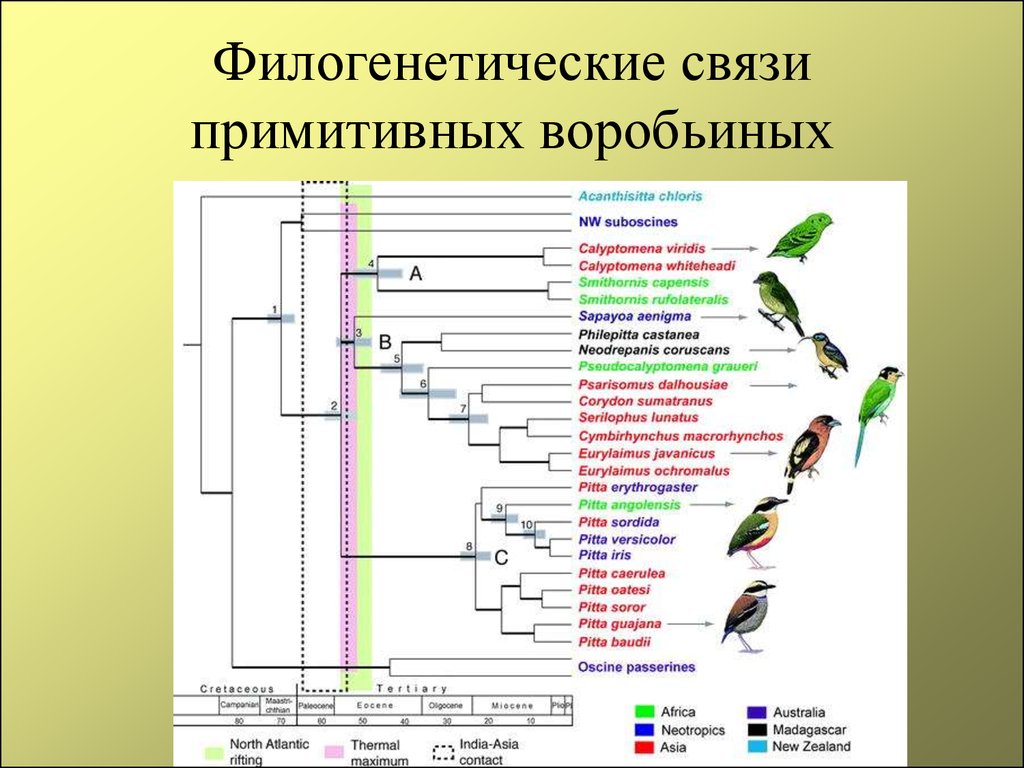 |
История версий
25.11.2018 — v1.4.4
Tracer больше не распространяется с этого веб-сайта.
Скомпилированные двоичные файлы (для Mac, Windows и Linux) доступны в репозитории FigTree GitHub.
2016-10-04 — v1.4.3
Исправление ошибок
Новые функции:
- Опция формы узла теперь может отображать формы для внутренних или внешних узлов или для обоих.
- Копирование названий выбранных таксонов, если они выбраны, поддерева, если выбраны ветви.
- Выбор «обратной оси» должен автоматически изменить масштабный коэффициент шкалы времени. Раньше пользователю нужно было установить это значение на -1.0.
- При поиске текста прокручивается, чтобы показать выделенную подсказку.
Исправлены ошибки:
- Проблема 102: Большие файлы SVG из figtree не работают
- Проблема 95: изменение исходного значения оси масштаба не работает
- Проблема 94: Легенда черты перекрывает дерево.
- Проблема 93: параметры экспорта PDF/SVG из командной строки не работают
- Проблема 92: Чтение файла с сочетанием целочисленных и реальных меток узлов вызывает исключение.
- Проблема 90: Изображение экспорта обрезает верхнюю часть самой высокой этикетки наконечника
- Проблема 79: экспорт .SVG создает поврежденные файлы
09.07.2014 — v1.4.2
Исправление ошибок
Новые функции:
- Новый параметр командной строки -url позволяет считывать деревья из URL-адресов в конвейерах.
Исправлены ошибки:
- Проблема 76: Ось шкалы должна отображать столько десятичных разрядов, сколько необходимо.
- Проблема 75: экспорт PNG и JPEG приводит к пустым изображениям.
- Проблема 64: Включение полос узлов переводит дерево вправо (теперь, я думаю, действительно исправлено).
14-06-2014 — v1.4.1
Исправления ошибок и новые функции
Новые функции:
- Копировать выбранные поддеревья в буфер обмена в формате NEXUS.
- Новые параметры экспорта графики (PDF, SVG, PNG и JPEG).
- Панель управления теперь прокручивается и может изменять размер.

Исправлены ошибки:
- Проблема 23: Панель поиска медленно открывается с большими деревьями.
- Проблема 28: фильтрация должна работать для текущих отображаемых меток.
- , проблема 53: Ярлыки подсказок по параметрам: «Цвет по» не отображает имена в цвете.
- Проблема 57: рутирование средней точки работает неправильно.
- Проблема 59: Clear Highlighting/Cartoon и т. д., похоже, не работает на отдельных ветвях.
- , проблема 62: когда все клады «свернуты», верхний треугольник обрезается.
- Проблема 64: При включении полос узла дерево смещается вправо.
- Проблема 69: Импорт аннотации вызывает сбой.
05.12.2012 — v1.4
Первая новая версия за 3 года!
Новые функции:
- Добавлен параметр «Форма узла», который может рисовать круги, квадраты или ромбы на каждом узле. Они могут быть окрашены и размер по атрибутам.
- Добавлен параметр «Легенда», который может создать цветовую легенду для любого используемого атрибута/цветовой схемы.
- Новая цветовая схема для дискретных атрибутов на основе цветового круга HSB. Это заменяет старую фиксированную последовательность цветов и более настраиваемо. Значения атрибутов могут быть переупорядочены.
- Новая цветовая схема для непрерывных атрибутов, также основанная на HSB. Старая цветовая схема интерполяции по-прежнему доступна.
- Цветовые схемы теперь можно выбирать для определенных атрибутов, они сохраняются и загружаются в файлы FigTree.
- Многочисленные изменения в элементах управления для большего удобства. К ним относятся повторное введение счетчиков размера шрифта (в дополнение к диалоговым окнам выбора шрифта) и прокрутка всей панели управления, когда она больше окна.
- Новое масштабирование/расширение. Максимальная степень масштабирования теперь пропорциональна количеству подсказок в дереве. Т.е. чем больше дерево, тем больше можно приближать. Также добавлена нелинейность масштабирования (начинается медленнее).
 Они могут быть окрашены и размер по атрибутам.
Они могут быть окрашены и размер по атрибутам. Т.е. чем больше дерево, тем больше можно приближать. Также добавлена нелинейность масштабирования (начинается медленнее).
Т.е. чем больше дерево, тем больше можно приближать. Также добавлена нелинейность масштабирования (начинается медленнее).Исправлены ошибки:
- Проблема 52: Спиннеры значительных фигур в настоящее время не имеют эффекта.
- , проблема 51: предпочтительные шрифты изначально установлены на первый шрифт в списке.
- Проблема 50: Проблема с корнем дерева MidPoint.
- Проблема 48: Метки таксонов не отображают признаки узлов.
- , проблема 45: средство импорта Nexus не игнорирует нераспознанные команды.
- , проблема 42: свернутые клады обрезаются в верхней части страницы.
- Проблема 39: повторное рутирование не всегда работает.
21-12-2009 — v1.3.1
Исправлены некоторые ошибки, появившиеся в последней версии
Новые функции: панель «Внешний вид», чтобы цветом каждой клады можно было управлять с помощью атрибута.
Исправлены ошибки:
- Повторно представлены графические форматы экспорта, которые были случайно опущены в предыдущей версии.
- Панель панели узлов теперь показывает доступные атрибуты для отображения (ошибка, появившаяся в предыдущей версии).
Ошибки, исправленные в v1.3
- Исправлена ошибка, из-за которой атрибуты «ветви» не загружались из файла NEXUS.
- Панель дерева теперь имеет фокус при открытии окна, что позволяет напрямую копировать/вставлять файлы дерева.
- На панели поиска доступны новые атрибуты.
- Типы аннотаций, определенные в диалоговом окне, не создавались.
14-12-2009 — v1.3
Новые функции раскраски, параметры экспорта и новый плагин QuickLook для Mac
Новые функции: ‘ панель, чтобы цветом в каждой кладе можно было управлять с помощью атрибута.
Исправлены ошибки:
- Исправлена ошибка, из-за которой атрибуты «ветви» не загружались из файла NEXUS.
- Панель дерева теперь имеет фокус при открытии окна, что позволяет напрямую копировать/вставлять файлы дерева.
- На панели поиска доступны новые атрибуты.
- Типы аннотаций, определенные в диалоговом окне, не создавались.
10.08.2009 — v1.2.3
Несколько новых функций и несколько исправлений ошибок
Новые функции:
- В настройки оси шкалы добавлена опция «Обратить ось» — она меняет направление оси шкалы на противоположное.
- Добавлена возможность включать и выключать линии сетки в настройках оси масштабирования.
Исправлены ошибки:
- Линии сетки шкалы могли не совпадать с осью.
- Типы аннотаций, определенные в диалоговом окне, не создавались.
24 февраля 2009 г. — v1.2.2
Несколько исправлений ошибок
Исправлены ошибки:
- Экспортированные деревья NEWICK теперь имеют конечную точку с запятой.
- Теперь текстовые поля на панелях управления снова доступны для редактирования.
- Полярные деревья с «Выравниванием меток кончиков» могут привести к тому, что дерево исчезнет со страницы.

09-01-2009 — v1.2.1
Несколько новых функций и несколько исправлений ошибок
Новые функции: установите шрифт для этих меток.
Исправлены ошибки:
- Исправлена проблема с перезагрузкой деревьев с целочисленными аннотациями.
- «Настройки» были недоступны в Windows и Linux (теперь они находятся в меню «Правка»).
- Теперь правильно экспортирует деревья «как показано» (т. е. укорененные или преобразованные).
30.11.2008 — v1.2
Несколько новых функций и множество исправлений ошибок
Новые функции:
- Добавлена кнопка «выделить», которая обеспечивает окраску блока для клады.
- Добавьте параметр непрерывного цветового градиента в элемент управления «Цвет по» на панели «Внешний вид». Используйте настройку для активации (в настоящее время она отключает параметр «Ширина по».
- Добавлена аннотация «Имя», которую можно использовать для редактирования названий таксонов. Его также можно использовать для именования кладов (и они отображаются для свернутых кладов).
- Добавлена новая раскрывающаяся панель поиска, которая активируется кнопкой «Найти» в меню «Правка» или значком поиска на панели инструментов.
- Размер шрифта и некоторые другие параметры теперь можно установить в окне «Настройки». Они действуют как значения по умолчанию для новых окон.
Исправлены ошибки:
- Исправлена проблема с применением аннотаций ветвей к неправильным ветвям при повторном укоренении дерева.
- Цвета «Цвет по» были изменены на противоположные по сравнению с цветами, предложенными в диалоговом окне «Настройка».
- Настройка «Цвет по» не работала должным образом, если автоматический выбор диапазона был отключен.
- Заголовки палитры элементов управления стали немного тоньше.
06.02.2008 — v1.1.2
Дополнительные форматы экспорта графики
Новые функции:
- Добавлены новые форматы экспорта графики с использованием библиотеки FreeHEP. Это включает
SVG, EPS, PDF и EMF, а также растровые форматы, такие как GIF, PNG и т. д. - Если дерево содержит «метки» (числа, такие как значения начальной загрузки перед
длина ветки), то пользователю предоставляется возможность дать им
информативное название.
Исправлены ошибки:
- Исправлено несколько мелких ошибок.
22-01-2008 — v1.1.1
Незначительное обновление исправляет несколько ошибок
Новые функции:
Римские цифры).
Палитра «Внешний вид».
 Они позволяют настраивать цвета и линии
Они позволяют настраивать цвета и линииширины.
горячие клавиши).
Исправлены ошибки:
- Фильтр (окно поиска на панели инструментов) не учитывал выбор
делается во всплывающем меню. - Параметры поиска и фильтрации практически не работали. Эти сейчас
работать по назначению. - FigTree аварийно завершает работу с исключением при запуске в некоторых вариантах Linux.
- Сбой FigTree при отображении меток узлов/ветвей после того, как дерево было
вручную повторно рутирован. - Варианты меток ветвей/узлов терялись при раскрашивании или инструментах аннотации
были использованы. - Удалены уродливые значки из пунктов меню, соответствующих панели инструментов.
кнопки.
13-01-2008 — v1.1
Основное обновление
Новые функции:
- Добавлены кнопки свертывания и мультипликации — сжатие создает один таксон, который представляет всю свернутую кладу, тогда как мультипликация создает треугольник который покрывает то же пространство, что и клада.